Termins "izaugsme", attiecinot to uz mikroorganismiem, nozīmē indivīda lieluma palielināšanos, bet "vairošanās" - indivīdu skaita palielināšanos populācijā. Pieaugot mikrobu šūnai, tās apjoms palielinās daudz straujāk nekā virsmas, tāpēc barības vielu izplatība šūnas citoplazmā kļūst mazāk efektīva un šūna dalās. Pirms tā sadalīšanas DNS molekulas tiek dublētas. Katra meitas šūna saņem mātes DNS kopiju.
Vienādos apstākļos audzētu dažādu mikrobu vairošanās ātrums ir atšķirīgs. Lielākajai daļai baktēriju ģenerācijas periodā (pagājušajā laikā) roorganismi var izmantot plašu oksidējamo organisko savienojumu klāstu, visbiežāk glikozi, no šiem savienojumiem enerģija tiek iegūta to oksidēšanās vai, precīzāk, elektronu ziedošanas rezultātā. viņiem.
Bioķīmisko procesu kopumu, kuru rezultātā izdalās šūnas dzīvībai nepieciešamā enerģija, sauc par elpošanu jeb bioloģisko oksidāciju. Saistībā ar mikroorganismiem viņi runā par anaerobiem un aerobiem elpošanas veidiem.
Starp divām secīgām šūnu dalīšanās reizēm) ir vidēji 15-30 minūtes; piemēram, Escherichia coli - 15-17 minūtes, vēdertīfa izraisītāji - 23 minūtes, korinebaktērijas difterija - 34 minūtes. Mycobacterium tuberculosis dalās lēnāk - reizi 18 stundās, spirohetes - 10 stundās.
Reprodukcijas metodes dažādās mikroorganismu grupās nav vienādas: baktērijas, riketsijas, spirohetas vairojas, daloties šķērsvirzienā divos līdzvērtīgos indivīdos. Grampozitīvās baktērijas sadalās, veidojot starpsienu, kas aug no perifērijas līdz centram. Mycobacterium tuberculosis gadījumā šūnas iekšpusē veidojas šķērseniskā starpsiena, pēc tam tā sadalās divos slāņos un šūna sadalās divās daļās.Starpsienas veidošanā piedalās gan citoplazmas membrāna, gan šūnas siena. Acīmredzot mezosoma, kas ir cieši saistīta ar citoplazmas membrānu, aktīvi piedalās baktēriju dalīšanās procesā. Gramnegatīvās baktērijas un riketsija centrā kļūst plānākas un tiek sadalītas divos indivīdos. Mezglu baktēriju un fraicisella reprodukcija notiek, veidojoties nierēm, kas pēc izmēra ir mazākas nekā sākotnējā šūna. Baktērijās notiek arī konjugācijas process - divu indivīdu pagaidu savienojums.
Baktēriju un spirohetu augšanu ne vienmēr pavada to sadalīšanās. Žults sāļi, ziepes, penicilīns, ultravioletie stari aizkavē šūnu dalīšanos, kā rezultātā veidojas gari pavedieni, kas ir daudz lielāki par sākotnējām šūnām.
Ievadot baktērijas uzturvielu barotnē, izšķir to augšanas un vairošanās fāzes, ko nosaka pieejamo barības avotu pieejamība un toksisko vielmaiņas produktu uzkrāšanās (21. att.).
Pirmā fāze - latentā (aizkavēšanās fāze) - atbilst baktēriju pielāgošanai jauniem eksistences apstākļiem. Šajā periodā baktērijas pielāgojas uzturvielu barotnei, to augšana netiek novērota.
Otrā fāze ir logaritmiskā augšana (eksponenciāla), kad baktērijas strauji aug, palielinās un, sasniedzot noteiktu izmēru, sāk dalīties divās meitas šūnās. Sadalīšanās šajā periodā notiek nemainīgā ātrumā. Katram baktēriju veidam vidējais paaudzes (vai dubultošanās) laiks ir atšķirīgs. Šajā laikā baktērijas ekstrahē barības vielas no barotnes, kā rezultātā tajā sāk uzkrāties vielmaiņas produkti.
Trešā fāze ir stacionāra augšana, kuras laikā organismu skaits kultūrā visu laiku paliek nemainīgs. Šajā periodā barības vielu daudzums barības vielu vidē ievērojami samazinās, un palielinās vielmaiņas produktu uzkrāšanās. Mikroorganismu dzīves apstākļi kļūst arvien mazāk labvēlīgi. Stacionārās fāzes ilgums dažādām baktērijām ir atšķirīgs.
Ceturtā fāze ir nāve, kad baktēriju šūnas kļūst mazākas un tās mirst. Šīs fāzes beigās mirstošo baktēriju skaits sāk dominēt pār dzīvotspējīgo šūnu skaitu. Pilnīga mikrobu nāve kultūrā var notikt dažu nedēļu vai mēnešu laikā atkarībā no mikroba veida, vides reakcijas un citiem faktoriem.
Vienkāršākie var vairoties ar šķērsenisko dalījumu, sašaurināšanos divos līdzvērtīgos indivīdos - amēbā un gareniskajā dalījumā - tripanosomās, žiardijās, balantīdijās. Pirms sadalīšanās divos indivīdos balantīdijas var apmainīties ar saviem kodoliem - mikrokodolus (konjugācijas process), malārijas plazmodijam ir aseksuālās un seksuālās attīstības cikls.
Vīrusi vairojas (vairojas) tikai saimniekorganisma dzīvās šūnās.
Vīrusa reprodukcijas process sastāv no vairākiem posmiem:
1) vīrusa iekļūšana šūnā;
2) intracelulārā reprodukcija;
3) vīrusa nobriešana un ārējo apvalku veidošanās dažiem vīrusiem; 4) vīrusa izolēšana no šūnas.
Vīrusa iekļūšanas process jutīgā šūnā sākas ar tā adsorbciju uz šūnas virsmas, kurai ir specifiski vīrusu receptori. Nukleīnskābju izdalīšanās process no kapsīda un ārējām membrānām sākas jau šūnas citoplazmas membrānā un beidzas citoplazmā (gripas vīruss, vaccinia).
Vīrusa intracelulārās reprodukcijas fāze jeb tā vairošanās parasti sākas ar šūnu makromolekulārās sintēzes nomākšanu. Visas šūnas enerģētiskās sistēmas, tās enzīmi, RNS, ribosomas sāk darboties vīrusa pavairošanai. Skartā šūna piegādā vīrusu ar nukleotīdiem nukleīnskābju veidošanai, aminoskābēm olbaltumvielām. Vīrusa RNS replikācija (angļu valodā replikācija - kopija, atkārtošana) tiek veikta ar enzīmu - polimerāžu palīdzību, un pati vīrusa RNS molekula kalpo kā matrica. DNS saturošajos vīrusos uz DNS šablona šūnas kodolā tiek sintezēta specifiska RNS, kas pēc tam nosaka vīrusa DNS un proteīna sintēzi. Vīrusu proteīni tiek sintezēti šūnas ribosomās.
Vīrusa daļiņas nobriešana, vīrusa nukleīnskābes noslēgšanās kapsīdā, notiek skartās šūnas kodolā (herpesvīrusi, adenovīrusi) vai citoplazmā (variola grupas vīrusi, rabdovīrusi, pikornavīrusi). Ārējo apvalku veidošanās miksovīrusos, togavīrusos notiek, izejot caur saimniekšūnas citoplazmas membrānu. Herpes vīruss saņem daļu no sava ārējā apvalka, izejot cauri šūnas kodola membrānai.
Vīrusa izolēšana no šūnas var notikt dažādos veidos. Miksovīrusus un togavīrusus, kad tie nobriest, šūna var izdalīties stundām ilgi, to nesabojājot. Poliomielīta vīruss (kuram nav ārējā apvalka) šūnā veidojas ātri, saglabājas tajā ilgu laiku un izdalās acumirklī, zibspuldzes veidā. Vīrusa un saimniekšūnas mijiedarbības galarezultāts var būt ātra šūnas iznīcināšana un nāve. Dažkārt vīrusi var atrasties šūnā ilgu laiku, neizraisot tās nāvi, un paliek bezgalīgā skaitā šūnu paaudžu – latentie vīrusi. Dažos gadījumos vīrusu var iznīcināt šūna bez redzamām sekām (abortīva vīrusu infekcija).
Baktēriju augšana un vairošanās
Baktēriju augšana notiek daudzu savstarpēji saistītu bioķīmisku reakciju rezultātā, kas sintezē šūnu materiālu, kā rezultātā palielinās visu ķīmisko komponentu daudzums. Baktērijās izšķir atsevišķu baktēriju šūnu augšanu un baktēriju augšanu populācijā.
individuāla baktēriju augšana. To vērtē pēc atsevišķu indivīdu lieluma pieauguma. Augšanas ātrums ir atkarīgs no ārējiem apstākļiem un pašas šūnas fizioloģiskā stāvokļa. Pastāvīgos apstākļos augšana notiek nemainīgā ātrumā. Stieņveida baktērijas pārsvarā aug garās ass virzienā, tāpēc šūnu augšanas laikā attiecība starp šūnas virsmu un tās tilpumu būtiski nemainās, un tas rada nemainīgus apstākļus katras šūnas daļas apgādāšanai ar barības vielām un skābekli. Cocci aug vienmērīgi visos virzienos, palielinot šūnas rādiusa lielumu, savukārt šūnas virsmas relatīvā vērtība samazinās, tāpēc piegādes apstākļi katrai šūnas daļai kļūst arvien nelabvēlīgāki. Intervālos starp šūnu dalīšanos baktērijas ir lielākas nekā tūlīt pēc dalīšanās.
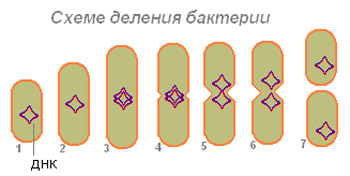
Baktēriju reprodukcija. Visbiežāk baktērijas vairojas binārās dalīšanās ceļā, kad no vienas šūnas veidojas divas šūnas, no kurām katra atkal dalās. Pirms dalīšanas procesa vienmēr notiek DNS replikācija. Ir divi dalīšanas veidi - sadalīšana ar sašaurināšanos (šņorēšana) un ar šķērseniskās starpsienas palīdzību (1.9. attēls).

1.9. attēls – baktēriju dalīšanās
A - sadalīšana ar sašaurināšanos; B - sadalīšana ar šķērssienu; CS, šūnu siena; CM, citoplazmas membrāna; H ir nukleoīds; P - sašaurināšanās
Sadalījums pēc sašaurinājuma(sasaukšanos) pavada šūnas sašaurināšanās tās dalīšanās vietā, un šajā procesā piedalās visi šūnu membrānu slāņi. Abās pusēs esošo membrānu izvirzījums šūnā to arvien vairāk sašaurina un, visbeidzot, sadala divās daļās. Šādi sadalās daudz gramnegatīvo baktēriju.
Sadalījums ar šķērseniskās starpsienas veidošanos raksturīgs grampozitīvām baktērijām. Tomēr dažās baktēriju grupās tika novērotas dalīšanas metožu izmaiņas (tioniskās baktērijas, mikobaktērijas). Sfēriskās baktērijās var veidoties vairākas šķērseniskas starpsienas (tetrakoki, sarkīni).
Laika posmu no dalīšanas līdz dalīšanai sauc šūnu cikls(baktēriju ontoģenēze). Ir vairāki veģetatīvā šūnu cikla veidi: monomorfs- veidojas tikai viena morfoloģiskā tipa šūnas (piemēram, baciļi), dimorfisks- divi morfoloģiskie veidi, polimorfs- vairākas, no kurām katrai ir raksturīgas noteiktas un nemainīgas šūnu cikla pazīmes (piemēram, aktinomicīti). Dimorfos un polimorfos ciklos izšķir meitas un mātes šūnas.
topošais Ubacterium ir binārās dalīšanās veids. Šī reprodukcijas metode ir raksturīga baktērijām, kurām ir dimorfs vai polimorfs šūnu cikls. Topošajām baktērijām ir šūnu polaritāte. Dažas baktērijas vairojas, izmantojot eksosporas (bet ne endosporas!), Hifa fragmentus (aktinomicītus). Ir baktērijas, kurām ir dzimumorgānu bārkstiņas, vai F-dzēra(angļu fertilit y-auglība, auglība), dzimuma faktora klātbūtnes dēļ.
Baktērijām ir raksturīgs augsts vairošanās ātrums. Piemēram, kad labvēlīgi apstākļi E. coli dalās ik pēc 20-30 minūtēm, 272 (72 paaudzes) mācīsies no vienas šūnas dienā. Apstākļos, kas izslēdz nāvi, šāda biomasa būs 4720 tonnas.Vairošanās ātrums ir atkarīgs no vides faktoriem (temperatūra, uztura apstākļi, mitrums, vides reakcija utt.) un no baktēriju sugas īpašībām. Augstais baktēriju vairošanās ātrums nodrošina to izdzīvošanu uz Zemes pat masveida nāves apstākļos. Atlikušās atsevišķās šūnas vairojas un atkal rada paaudzi.
Baktēriju augšana populācijā. Populācija (fr. populācija - populācija) ir vienas sugas baktēriju kopums (tīrkultūra) vai dažādi veidi(jaukta asociācija), attīstās ierobežotā telpā (piemēram, uzturvielu vidē). Baktēriju populācijā šūnas nepārtraukti aug, vairojas un mirst. Mikroorganismu audzēšana mākslīgos apstākļos ir periodiska, nepārtraukta un sinhrona.
Periodiska (stacionāra) audzēšana. Šī kultivēšana notiek bez barības barotnes pieplūdes un aizplūšanas. To raksturo klasika mikroorganismu augšanas līkne, kurās izšķir atsevišķas baktēriju populācijas augšanas fāzes, atspoguļojot vispārējs modelisšūnu augšana un vairošanās (1.10. attēls).

1.10. attēls – baktēriju populācijas augšanas un attīstības līkne
Kavēšanās fāze(angļu val. lag — lag) sākas no brīža, kad baktērijas tiek stādītas svaigā barotnē. Šūnas pielāgojas dotajiem kultivēšanas apstākļiem, aug, bet nevairojas, sasniedz maksimālo augšanas ātrumu. Absolūtais un īpatnējais augšanas ātrums palielinās no nulles līdz maksimāli iespējamajām vērtībām.
Absolūtais izaugsmes temps nosaka attiecība:
V = dx/dt, (1,1)
kur V ir biomasas vai šūnu skaita pieaugums;.
x ir biomasa vai šūnu skaits,
t ir laiks.
Īpatnējais augšanas ātrums nosaka pēc formulas:
µ = (dx/dt) ∙ 1/х, (1,2)
kur µ ir biomasas pieaugums e laika vienībā uz biomasas vienību,
x ir sākotnējā biomasa.
Kavēšanās fāzes ilgums ir atkarīgs no baktēriju bioloģiskajām īpašībām, kultūras vecuma, inokulāta daudzuma, barotnes sastāva, temperatūras, aerācijas, pH utt. Dažām baktērijām ir īss augšanas aizkavēšanās periods. , savukārt citiem ir garš. Jo jaunāka kultūra, jo īsāks periods. Jo tuvāk barotnes sastāvs tam, kurā mikroorganismi tika audzēti, jo īsāka ir nobīdes fāze. Barības barotnes izmaiņas izraisa izmaiņas aizkavēšanās fāzē, jo fermentu sintēzei vai to aktivitātes palielināšanai ir nepieciešams laiks. Tādējādi augšanas aizkavēšanās faktorus var iedalīt ārējā(vidējais sastāvs, pH, temperatūra utt.) un iekšējais(kultūras laikmets). Fāzes ilgums var būt no vairākām minūtēm līdz vairākām stundām un pat dienām. Šajā fāzē μ = 0 .
logaritmisks, vai eksponenciāls, vai log fāze, ko raksturo maksimālais baktēriju dalīšanās ātrums. Eksponenciālo iedzīvotāju skaita pieaugumu apraksta ar vienādojumu:
X \u003d X o ∙ e μ max ∙ t, (1.3)
kur Chi X o - šūnu (vai biomasas) skaits eksperimenta beigās un sākumā;
t ir eksperimenta laiks;
e ir naturālā logaritma bāze;
µmaks – maksimālais īpatnējais augšanas ātrums.
Logaritmiskās fāzes laikā lielākā daļa šūnu ir fizioloģiski jaunas, bioķīmiski aktīvas un arī visjutīgākās pret nelabvēlīgiem vides faktoriem. Šajā fāzē μ = max Šī fāze ir daudzpakāpju, jo tās augšanas sākumā baktērijas aug vidē ar substrāta pārpalikumu, tad samazinās to koncentrācija, mainās enzīmu aktivitāte un šūnu saturs. metabolīti palielinās. Turklāt baktēriju vairošanos ietekmē daudzi faktori: baktēriju sugu īpašības, barības barotnes raksturs un tās atsevišķo komponentu koncentrācija, kā arī kultivēšanas temperatūra.
Lēnas augšanas fāze. Tas apvieno divas fāzes - lineārās augšanas fāze(μ = const) un negatīvā paātrinājuma fāze. Fāzei lineārās augšanas periodā ir raksturīgs nemainīgs biomasas pieauguma ātrums (šūnu skaits). Tad, pārejot uz negatīvā paātrinājuma fāzi, sadalošo šūnu skaits samazinās. Fāzes iestāšanās ir skaidrojama ar kvantitatīvajām izmaiņām uzturvielu barotnes sastāvā (barības vielu patēriņš, vielmaiņas produktu uzkrāšanās).
Stacionārā fāze ko raksturo līdzsvars starp mirstošām un jaunizveidotajām šūnām. Faktori, kas ierobežo baktēriju augšanu iepriekšējā fāzē, ir stacionārās fāzes cēlonis. Biomasa nepalielinās (μ = 0) Šajā fāzē tiek novērota maksimālā biomasas vērtība un maksimālais kopējais šūnu skaits. Šīs maksimālās vērtības tiek sauktas ražu, vai izeja. Viens no ierobežojošajiem faktoriem ir maksimālā šūnu koncentrācija uz barības barotnes tilpuma vienību. Dažādu veidu baktērijās šī vērtība ievērojami atšķiras. Stacionārajā fāzē šūnām raksturīga nelīdzsvarota augšana (šūnu komponenti tiek sintezēti dažādos ātrumos), vielmaiņas procesu intensitātes samazināšanās un lielāka izturība pret fizikālām un ķīmiskām ietekmēm.
mirstošā fāze (eksponenciāla šūnu nāve) raksturo dzīvo šūnu skaita samazināšanās, populācijas neviendabīguma palielināšanās (parādās šūnas, kas neuztver krāsvielu, ar vāju mureīna slāņa attīstību utt.). Miršanas process dominē pār dalīšanu (μ< 0).
Izdzīvošanas fāze ko raksturo atsevišķu šūnu klātbūtne, kas ilgu laiku ir saglabājušas dzīvotspēju vairuma šūnu nāves apstākļos. Izdzīvojušajām šūnām raksturīga zema vielmaiņas procesu aktivitāte, izmaiņas šūnu ultrastruktūrā (smalkgraudaina citoplazma, poliribosomu trūkums utt.). Šūnas ir izturīgākas pret nelabvēlīgiem vides apstākļiem.
Tādējādi stacionārās kultivēšanas laikā mikrobu šūnas pastāvīgi atrodas mainīgos apstākļos: vispirms visu barības vielu ir pārpalikums, tad pamazām iestājas to trūkums, tad šūnas tiek saindētas ar vielmaiņas produktiem.
Ierobežojošo faktoru ietekme uz izaugsmes tempu. Normālai mikroorganismu augšanai un attīstībai vidē jāsatur nepieciešamās barības vielas, jābūt atbilstošam pH, temperatūrai utt. Tiek saukti faktori, kas ierobežo ražas augšanu ierobežojoši. Mikroorganismu populācijas augšanas raksturīga iezīme ir īpatnējā augšanas ātruma atkarība no substrāta koncentrācijas. Šī atkarība ir izteikta Monoda vienādojums, kas ir hiperboliska funkcija:
μ = μ max ∙ S/(S + K S), (1,4)
kur μ ir īpatnējais augšanas ātrums;
μ max - maksimālais īpatnējais augšanas ātrums;
S ir substrāta koncentrācija;
K S - piesātinājuma konstante, skaitliski vienāda ar tādu substrāta koncentrāciju, kas nodrošina augšanas ātrumu, kas atbilst pusei no μ max vērtības.
Patērējot barības vielas, vide tiek bagātināta ar vielmaiņas produktiem, kas arī ierobežo kultūras augšanu. Vispārīgākais gadījums par substrāta un vielmaiņas produktu koncentrācijas ietekmi uz mikroorganismu populācijas augšanas ātrumu tika atspoguļots N.D. Jeruzalemes modelī:
μ = μ max ∙ S/(S + K S) ∙ K P / (K P / + P), (1,5)
kur P ir vielmaiņas produktu koncentrācija;
K P ir konstante, kas skaitliski vienāda ar tādu vielmaiņas produktu koncentrāciju, pie kuras augšanas ātrums palēninās uz pusi.
Šī vienādojuma analīze parāda, ka pie nosacījuma K P >> P, kad P vērtību var neņemt vērā. augšanas ātrumu ierobežo tikai substrāta koncentrācija. Ja S >> K S , tad augšanas ātrumu ierobežo vielmaiņas produktu uzkrāšanās
Nepārtraukta audzēšana. Ja tvertnē, kurā atrodas baktēriju populācija, nepārtraukti tiek piegādāta svaiga barotne un vienlaikus ar tādu pašu ātrumu tiek izņemts baktēriju šūnas un vielmaiņas produktus saturošais kultūras šķidrums, tad tiek iegūta nepārtraukta kultivēšana. Regulējot plūsmas vides ātrumu, baktēriju populācijas augšanu var kontrolēt, piemēram, lai pagarinātu logaritmisko vai stacionāro fāzi uz jebkuru vēlamo laiku. Nepārtraukta kultivēšana tiek veikta īpašās ierīcēs - chemostatos un turbidostatos.
Ķīmostāti. Baktēriju augšanu regulē substrāta koncentrācija. Uzturot nemainīgu viena no nepieciešamajiem substrātiem (slāpekļa vai oglekļa avota) koncentrāciju, regulējot barotnes plūsmas ātrumu, ir iespējams līdzsvarot kultūras augšanas ātrumu. Šūnu biomasas vērtības izmaiņu ātrums chemostatā ir vienāds ar starpību starp biomasas augšanas ātrumu un tās izņemšanas ātrumu no kultivatora. Iedzīvotāju blīvums paliek nemainīgs, ja μ=D (īpatnējais augšanas ātrums ir vienāds ar atšķaidīšanas koeficientu), t.i. tiek līdzsvarots šūnu zudums izskalošanās rezultātā un to pieaugums vairošanās rezultātā.
Turbidostats.Darbības principa pamatā ir vides plūsmas ātruma regulēšana pēc iedzīvotāju blīvuma. Iedzīvotāju blīvumu kontrolē fotoelements, kas savienots ar releju, kas regulē barotnes plūsmu. Kad iedzīvotāju blīvums sasniedz iepriekš noteiktu līmeni, tiek aktivizēts relejs un kultivatorā nonāk svaiga barotne. Rezultātā šūnu koncentrācija samazinās līdz noteiktam līmenim, un tad barotnes padeve tiek automātiski izslēgta.
Turbidostatiskā kontrole var būt balstīta uz citām metodēm biomasas vai produktu, kas veidojas baktēriju augšanas laikā, noteikšanai (piemēram, pH-stat metode plūsmas ātruma regulēšanai, oksistata lietošana - plūsmas ātruma kontrole pēc skābekļa patēriņa ātruma utt.).
Nepārtraukta mikroorganismu kultivēšana tiek izmantota, lai pētītu to fizioloģiju, bioķīmiju, ģenētiku u.c., un to plaši izmanto arī mikrobioloģiskajā rūpniecībā.
Sinhronā audzēšana . Sinhronās kultūras ir kultūras, kurās kādu laiku visas šūnas dalās vienlaicīgi (sinhroni) visu indivīdu vienādas gatavības dalīties dēļ. Sinhronizācija tiek panākta ar fizikālām un ķīmiski bioloģiskām metodēm. Fizikālās metodes ir temperatūras ietekme, diferenciālā centrifugēšana vai diferenciālā filtrēšana utt. Ķīmiskās un bioloģiskās metodes: baktēriju piespiedu badošanās, baktēriju kultivēšana uz zemākām barotnēm ar to tālāku pārvietošanu uz pilnu barotni. Sinhronās kultūras izmanto ģenētiskai un citoloģiskie pētījumi, pētīt atsevišķu šūnu komponentu sintēzi baktēriju dalīšanās procesā.
Baktēriju augšana notiek daudzu savstarpēji saistītu bioķīmisku reakciju rezultātā, kas veic šūnu materiāla sintēzi. Baktērijās izšķir atsevišķu baktēriju šūnu augšanu un baktēriju augšanu populācijā.
Individuālo izaugsmi vērtē pēc atsevišķu indivīdu lieluma pieauguma. Augšanas ātrums ir atkarīgs no ārējiem apstākļiem un pašas šūnas fizioloģiskā stāvokļa. Pastāvīgos apstākļos augšana notiek nemainīgā ātrumā. Stieņveida baktērijas pārsvarā aug garās ass virzienā, koki aug vienmērīgi visos virzienos. Intervālā starp šūnu dalīšanos baktērijas ir lielākas nekā tūlīt pēc dalīšanās.
Baktēriju reprodukcija
Visbiežāk baktērijas vairojas binārās dalīšanās ceļā, kad no vienas šūnas veidojas divas šūnas, no kurām katra atkal dalās. Pirms dalīšanas procesa vienmēr notiek DNS replikācija (dubultošanās). Ir divi dalīšanas veidi - sadalīšana ar sašaurināšanos (šņorēšana) un ar šķērseniskās starpsienas palīdzību (A.7. attēls).
Sadalījums pēc sašaurinājuma(sasaukšanos) pavada šūnas sašaurināšanās tās dalīšanās vietā, un šajā procesā piedalās visi šūnu membrānu slāņi. Membrānu izvirzījums šūnā to arvien vairāk sašaurina un, visbeidzot, sadala divās daļās. Šis sadalījums ir raksturīgs gramnegatīvām baktērijām. Sadalījums ar šķērseniskās starpsienas veidošanos raksturīgs grampozitīvām baktērijām. Tomēr dažās baktēriju grupās tika novērotas dalīšanas metožu izmaiņas (tioniskās baktērijas, mikobaktērijas). Sfēriskās baktērijās var veidoties vairākas šķērseniskas starpsienas (tetrakoki, sarkīni). topošais Ubacterium ir binārās dalīšanās veids. Šī reprodukcijas metode ir raksturīga baktērijām, kurām ir dimorfs vai polimorfs šūnu cikls. Topošajām baktērijām ir šūnu polaritāte. Dažas baktērijas vairojas ar eksosporu (bet ne endosporu!) palīdzību, dažas – ar hifu (aktinomicītu) fragmentiem. Dažām baktērijām ir dzimumorgānu bārkstiņi, vai F-dzēra.
Laika posmu no dalīšanas līdz dalīšanai sauc šūnu cikls. Ir vairāki veģetatīvā šūnu cikla veidi: monomorfs- veidojas tikai viena morfoloģiskā tipa šūnas (piemēram, baciļi), dimorfisks- divi morfoloģiskie šūnu veidi, polimorfs- vairāki (actinomycetes). Dimorfos un polimorfos ciklos izšķir meitas un mātes šūnas.
Baktērijām ir raksturīgs augsts vairošanās ātrums. Piemēram, labvēlīgos apstākļos Escherichia coli dalās ik pēc 20-30 minūtēm, tas dod 2 72 dienā, t.i. 72 paaudzes. Apstākļos, izņemot nāvi, šī biomasa būs 4720 tonnas.Vairošanās ātrums ir atkarīgs no vides faktoriem (temperatūra, uztura apstākļi, mitrums, vides reakcija utt.) un no baktēriju sugas īpašībām. Augstais baktēriju vairošanās ātrums nodrošina to izdzīvošanu uz zemes pat masveida nāves apstākļos. Atlikušās atsevišķās šūnas vairojas un atkal rada paaudzi.
Baktēriju augšana populācijā. iedzīvotāju (fr. populācija- populācija) ir vienas sugas (tīrkultūra) vai dažādu sugu (jaukta asociācija) baktēriju kolekcija, kas attīstās ierobežotā telpā (piemēram, barotnē). Baktēriju populācijā šūnas nepārtraukti aug, vairojas un mirst. Mikroorganismu audzēšana mākslīgos apstākļos ir periodiska, nepārtraukta un sinhrona.
Periodiska (stacionāra) audzēšana notiek bez barības barotnes pieplūdes un aizplūšanas. To raksturo klasika mikroorganismu augšanas līkne, kurā izšķir atsevišķas baktēriju populācijas augšanas fāzes, kas atspoguļo vispārējo šūnu augšanas un vairošanās modeli.
Kavēšanās fāze(Angļu) aizkavēšanās- atpalicība) sākas no brīža, kad baktērijas tiek sētas svaigā barotnē. Šūnas pielāgojas dotajiem kultivēšanas apstākļiem, aug, bet nevairojas, sasniedz maksimālo augšanas ātrumu. Absolūtais un īpatnējais augšanas ātrums palielinās no nulles līdz maksimāli iespējamajām vērtībām.
Absolūtais izaugsmes temps nosaka attiecība:
V = dx/dt , (1.1)
kur V - biomasas vai šūnu skaita pieaugums, kas izteikts masas vienībās, šūnu skaits vai parastās vienības laika vienībā.
X- biomasa vai šūnu skaits;
t- laiks.
Īpatnējais augšanas ātrums nosaka pēc formulas:
µ = (dx/dt) ? 1/x, (1.2)
kur µ - biomasas pieaugums e laika vienība uz biomasas vienību,
X- sākotnējā biomasa.
Kavēšanās fāzes ilgums ir atkarīgs no baktēriju bioloģiskajām īpašībām, kultūras vecuma, inokulāta daudzuma, barotnes sastāva, temperatūras, aerācijas, pH utt. Dažām baktērijām ir īss augšanas aizkavēšanās periods. , savukārt citiem ir garš. Jo jaunāka kultūra, jo īsāks periods. Jo tuvāk barotnes sastāvs tam, kurā mikroorganismi tika audzēti, jo īsāka ir nobīdes fāze. Barības barotnes izmaiņas izraisa izmaiņas aizkavēšanās fāzē, jo fermentu sintēzei vai to aktivitātes palielināšanai ir nepieciešams laiks. Tādējādi augšanas aizkavēšanās faktorus var iedalīt ārējos (vidējais sastāvs, pH, temperatūra utt.) un iekšējos (kultūras vecums). Fāzes ilgums var būt no vairākām minūtēm līdz vairākām stundām un pat dienām. Šajā fāzē μ = 0 .
log fāze (logaritmisks, vai eksponenciāls) raksturo maksimālais baktēriju dalīšanās ātrums. Kopējo baktēriju skaitu nosaka pēc formulas:
N = N 0 = 2 n, (1.3)
kur N un N0- kopējais šūnu skaits attiecīgi fāzes beigās un fāzes sākumā;
n ir paaudžu vai paaudžu skaits.
Mikrobioloģiskajā praksē izteikt kopējais skaits mikrobu šūnas visbiežāk izmanto nevis absolūtos skaitļus (jo tie sasniedz milzīgas vērtības), bet gan to logaritmus. Pēc (1.3) vienādojuma logaritma ņemšanas: lg N = lg N 0 + n?lg2, n?lg2 = lg N - lg N 0, tātad paaudžu skaits ir: n = (lg N - lg N 0)/ lg2
Vienas šūnas vairošanās ātrums vai paaudzes periods:
g = t/n , (1.4)
kur t - laiks;
n - paaudžu skaits;
g- paaudzes periods.
Līdzekļi: g \u003d t ?lg 2 / (lg N - lg N 0) (1.5)
Iepriekš minētie vienādojumi ir balstīti uz pieņēmumu, ka log fāzē visas 100% šūnu ir dzīvotspējīgas. Taču eksperimentāli noskaidrots, ka pat šajā fāzē iet bojā ap 20% šūnu, tāpēc iepriekš minētajās formulās tiek veikts grozījums - 2 vietā tiek ņemts 1,6.
Eksponenciālo iedzīvotāju skaita pieaugumu apraksta ar vienādojumu:
X \u003d X 0? e μ max ? t , (1.6)
kur X un X 0 - šūnu (vai biomasas) skaits attiecīgi eksperimenta beigās un sākumā;
t- pieredzes laiks;
e ir naturālā logaritma bāze;
µmax - maksimālais īpatnējais augšanas ātrums.
Logaritmiskās fāzes laikā lielākā daļa šūnu ir fizioloģiski jaunas, bioķīmiski aktīvas un arī visjutīgākās pret nelabvēlīgiem vides faktoriem. Šajā fāzē μ = maks .
Lēnas augšanas fāze. Tas apvieno divas fāzes - lineārās augšanas fāze(μ = const) un negatīvā paātrinājuma fāze. Fāzei lineārās augšanas periodā ir raksturīgs nemainīgs biomasas pieauguma ātrums (šūnu skaits). Tad, pārejot uz negatīvā paātrinājuma fāzi, sadalošo šūnu skaits samazinās. Fāzes iestāšanās ir skaidrojama ar kvantitatīvajām izmaiņām uzturvielu barotnes sastāvā (barības vielu patēriņš, vielmaiņas produktu uzkrāšanās).
Stacionārā fāze ko raksturo līdzsvars starp mirstošām un jaunizveidotajām šūnām. Faktori, kas ierobežo baktēriju augšanu iepriekšējā fāzē, ir stacionārās fāzes cēlonis. Nav biomasas pieauguma (μ = 0 ) . Šajā fāzē tiek novērota maksimālā biomasas vērtība un maksimālais kopējais šūnu skaits. Šīs maksimālās vērtības tiek sauktas ražu, vai izeja.
mirstošā fāze (eksponenciāla šūnu nāve) raksturo dzīvo šūnu skaita samazināšanās, populācijas neviendabīguma palielināšanās (parādās šūnas, kas neuztver krāsvielu, ar vāju mureīna slāņa attīstību utt.). Miršanas process dominē pār dalīšanu (μ< 0).
Izdzīvošanas fāze ko raksturo atsevišķu šūnu klātbūtne, kas ilgu laiku ir saglabājušas dzīvotspēju vairuma šūnu nāves apstākļos. Izdzīvojušajām šūnām raksturīga zema vielmaiņas procesu aktivitāte, izmaiņas šūnu ultrastruktūrā (smalkgraudaina citoplazma, poliribosomu trūkums utt.). Šūnas ir izturīgākas pret nelabvēlīgiem vides apstākļiem.
Tādējādi stacionārās kultivēšanas laikā mikrobu šūnas pastāvīgi atrodas mainīgos apstākļos: vispirms visu barības vielu ir pārpalikums, tad pamazām iestājas to trūkums, tad šūnas tiek saindētas ar vielmaiņas produktiem.
Ierobežojošo faktoru ietekme uz izaugsmes tempu. Normālai mikroorganismu augšanai un attīstībai vidē jāsatur nepieciešamās barības vielas, jābūt atbilstošam pH, temperatūrai utt. Tiek saukti faktori, kas ierobežo ražas augšanu ierobežojoši. Mikroorganismu populācijas augšanas raksturīga iezīme ir īpatnējā augšanas ātruma atkarība no substrāta koncentrācijas. Šī atkarība ir izteikta Monoda vienādojums, kas ir hiperboliska funkcija:
μ = μmax? S/(S + K S), (1.7)
kur μ - īpatnējais augšanas ātrums ;
µmaks- maksimālais īpatnējais augšanas ātrums;
S- substrāta koncentrācija;
K S- piesātinājuma konstante, skaitliski vienāda ar tādu substrāta koncentrāciju, kas nodrošina augšanas ātrumu, kas atbilst pusei no μ max vērtības.
Patērējot barības vielas, vide tiek bagātināta ar vielmaiņas produktiem, kas arī ierobežo kultūras augšanu. Vispārīgākais gadījums substrāta un vielmaiņas produktu koncentrācijas ietekmei uz mikroorganismu populācijas augšanas ātrumu tika atspoguļots N.D. modelī. Jeruzaleme:
μ = μmax? S/(S + K S) ? K P / (K P + P), (1.8)
kur R- vielmaiņas produktu koncentrācija;
K R- konstante skaitliski vienāda ar tādu vielmaiņas produktu koncentrāciju, pie kuras augšanas ātrums palēninās uz pusi.
Šī vienādojuma analīze parāda, ka pie nosacījuma K P >> P, kad P vērtību var neņemt vērā. augšanas ātrumu ierobežo tikai substrāta koncentrācija. Ja S >> K S , tad augšanas ātrumu ierobežo vielmaiņas produktu uzkrāšanās.
Nepārtraukta audzēšana. Ja tvertnē, kurā atrodas baktēriju populācija, nepārtraukti tiek piegādāta svaiga barotne un vienlaikus ar tādu pašu ātrumu tiek izņemts baktēriju šūnas un vielmaiņas produktus saturošais kultūras šķidrums, tad tiek iegūta nepārtraukta kultivēšana. Regulējot plūsmas vides ātrumu, baktēriju populācijas augšanu var kontrolēt, piemēram, lai pagarinātu logaritmisko vai stacionāro fāzi uz jebkuru vēlamo laiku. Nepārtraukta kultivēšana tiek veikta īpašās ierīcēs - chemostatos un turbidostatos. Nepārtraukta mikroorganismu kultivēšana tiek izmantota, lai pētītu to fizioloģiju, bioķīmiju, ģenētiku u.c., un to plaši izmanto arī mikrobioloģiskajā rūpniecībā.
Sinhronās kultūras- tās ir kultūras, kurās kādu laiku visas šūnas dalās vienlaicīgi (sinhroni), pateicoties vienādai gatavībai visu indivīdu dalīšanai. Sinhronizācija tiek panākta ar fizikālām un ķīmiski bioloģiskām metodēm. Fizikālās metodes ietver temperatūras pakļaušanu, diferenciālo centrifugēšanu vai diferenciālo filtrēšanu, ķīmiski bioloģiskās metodes - baktēriju piespiedu badu, baktēriju kultivēšanu uz zemākām barotnēm ar to turpmāku pārvietošanu uz pilnu barotni. Sinhronās kultūras izmanto ģenētiskiem un citoloģiskiem pētījumiem, atsevišķu šūnu komponentu sintēzes pētīšanai baktēriju dalīšanās procesā.
Baktēriju pavairošana dalīšanās ceļā ir visizplatītākā mikrobu populācijas palielināšanas metode. Pēc dalīšanās baktērijas izaug līdz sākotnējam izmēram, kam nepieciešamas noteiktas vielas (augšanas faktori).
Baktēriju pavairošanas metodes ir atšķirīgas, taču lielākajai daļai to sugu aseksuālas vairošanās veids ir raksturīgs dalīšanas metodei. Baktērijas reti vairojas, veidojot pumpurus. Baktēriju seksuālā pavairošana notiek primitīvā formā.
Rīsi. 1. Fotoattēlā baktēriju šūna atrodas dalīšanās stadijā.
Baktēriju ģenētiskais aparāts
Baktēriju ģenētisko aparātu pārstāv viena DNS - hromosoma. DNS ir noslēgta gredzenā. Hromosoma atrodas nukleotīdā, kuram nav membrānas. Baktēriju šūna satur plazmīdas.
Nukleoīds
Nukleoīds ir analogs kodolam. Tas atrodas šūnas centrā. Tajā ir lokalizēta DNS - iedzimtas informācijas nesējs salocītā veidā. Nesavītā DNS garums sasniedz 1 mm. Baktēriju šūnas kodolvielai nav membrānas, kodola un hromosomu kopas, un to nedala mitoze. Pirms dalīšanas nukleotīds tiek dubultots. Dalīšanās laikā nukleotīdu skaits palielinās līdz 4.

Rīsi. 2. Fotoattēlā baktēriju šūna uz griezuma. Centrālajā daļā ir redzams nukleotīds.
Plazmīdas
Plazmīdas ir autonomas molekulas, kas salocītas divpavedienu DNS gredzenā. To masa ir daudz mazāka par nukleotīda masu. Neskatoties uz to, ka iedzimtā informācija ir iekodēta plazmīdu DNS, tās nav vitāli svarīgas un nepieciešamas baktēriju šūnai.

Rīsi. 3. Fotoattēlā redzama baktēriju plazmīda.
Sadalīšanas posmi
Pēc noteikta izmēra sasniegšanas, kas raksturīga pieaugušai šūnai, tiek iedarbināti dalīšanās mehānismi.
DNS replikācija
DNS replikācija notiek pirms šūnu dalīšanās. Mezosomas (citoplazmas membrānas krokas) aiztur DNS, līdz dalīšanās (replikācijas) process ir pabeigts.
DNS replikācija tiek veikta ar DNS polimerāzes enzīmu palīdzību. Replikācijas laikā tiek pārrautas ūdeņraža saites 2-pavedienu DNS, kā rezultātā no vienas DNS veidojas divas meitas vienpavediena. Pēc tam, kad meitas DNS ir ieņēmusi savu vietu atdalītajās meitas šūnās, tās tiek atjaunotas.
Tiklīdz DNS replikācija ir pabeigta, sintēzes rezultātā parādās sašaurināšanās, kas sadala šūnu uz pusēm. Pirmkārt, sadalās nukleotīds, pēc tam citoplazma. Šūnu sieniņu sintēze pabeidz dalīšanos.
Rīsi. 4. Baktēriju šūnu dalīšanās shēma.
DNS segmentu apmaiņa
Siena baciļos DNS replikācijas process tiek pabeigts, apmainoties ar 2 DNS segmentiem.
Pēc šūnu dalīšanās veidojas tilts, pa kuru vienas šūnas DNS pāriet citā. Pēc tam abas DNS savijas. Daži abu DNS posmi salīp kopā. Adhēzijas vietās notiek DNS segmentu apmaiņa. Viena no DNS atgriežas pirmajā šūnā gar džemperi.

Rīsi. 5. DNS apmaiņas variants siena bacilī.
Baktēriju šūnu dalīšanās veidi
Ja šūnu dalīšanās ir pirms dalīšanās procesa, tad veidojas daudzšūnu stieņi un koki.
Ar sinhronu šūnu dalīšanos veidojas divas pilnvērtīgas meitas šūnas.
Ja nukleotīds dalās ātrāk nekā pati šūna, tad veidojas daudznukleotīdu baktērijas.
Baktēriju atdalīšanas veidi
Sadalīšana laužot
Sadalīšanās ar laušanu ir raksturīga Sibīrijas mēra baciļiem. Šīs dalīšanās rezultātā šūnas saplīst locītavās, pārraujot citoplazmas tiltus. Tad viņi viens otru atgrūž, veidojot ķēdes.
bīdāmā atdalīšana
Ar bīdāmu atdalīšanu pēc dalīšanas šūna atdalās un it kā slīd pa citas šūnas virsmu. Šī atdalīšanas metode ir raksturīga dažām Escherichia formām.
sadalīt sadalīt
Ar dalītu dalījumu viena no sadalītajām šūnām apraksta apļa loku ar tā brīvo galu, kura centrs ir tās saskares punkts ar citu šūnu, veidojot romiešu pieci jeb ķīļrakstu (corynebacterium diphtheria, listeria).

Rīsi. 6. Fotoattēlā stieņveida baktērijas, kas veido ķēdes (sibīrijas mēra nūjiņas).

Rīsi. 7. Fotoattēlā bīdāmā metode Escherichia coli atdalīšanai.

Rīsi. 8. Sadalīšanas metode korinebaktēriju atdalīšanai.
Baktēriju kopu skats pēc dalīšanās
Sadalošo šūnu uzkrājumiem ir dažādas formas, kas ir atkarīgas no dalīšanas plaknes virziena.
lodveida baktērijas sakārtoti pa vienam, pa diviem (diplokoki), maisos, ķēdēs vai kā vīnogu ķekarus. Stieņveida baktērijas - ķēdēs.
Spirālveida baktērijas- haotisks.

Rīsi. 9. Fotoattēlā redzami mikrokoki. Tie ir apaļi, gludi, baltā, dzeltenā un sarkanā krāsā. Mikrokoki dabā ir visuresoši. Viņi dzīvo dažādos cilvēka ķermeņa dobumos.

Rīsi. 10. Fotoattēlā diplokoku baktērijas - Streptococcus pneumoniae.

Rīsi. 11. Fotoattēlā Sarcina baktērijas. Coccoid baktērijas tiek apvienotas paciņās.

Rīsi. 12. Fotoattēlā streptokoku baktērijas (no grieķu "streptos" - ķēde). Sakārtotas ķēdēs. Tie ir vairāku slimību izraisītāji.

Rīsi. 13. Fotoattēlā baktērijas ir "zelta" stafilokoki. Sakārtota kā "vīnogu ķekars". Klasteriem ir zelta krāsa. Tie ir vairāku slimību izraisītāji.

Rīsi. 14. Fotoattēlā leptospira vītņotās baktērijas ir daudzu slimību izraisītāji.

Rīsi. 15. Fotoattēlā Vibrio ģints nūjiņveida baktērijas.
baktēriju dalīšanās ātrums
Baktēriju dalīšanās ātrums ir ārkārtīgi augsts. Vidēji viena baktēriju šūna dalās ik pēc 20 minūtēm. Tikai vienas dienas laikā viena šūna veido 72 pēcnācēju paaudzes. Mycobacterium tuberculosis sadalās lēni. Viss sadalīšanas process viņiem aizņem apmēram 14 stundas.

Rīsi. 16. Fotoattēlā redzams streptokoku šūnu dalīšanās process.
Baktēriju seksuālā pavairošana
1946. gadā zinātnieki atklāja seksuālo reprodukciju primitīvā formā. Šajā gadījumā gametas (vīriešu un sieviešu dzimumšūnas) neveidojas, tomēr dažas šūnas apmainās ar ģenētisko materiālu ( ģenētiskā rekombinācija).
Gēnu pārnešana notiek kā rezultātā konjugācijas— ģenētiskās informācijas daļas vienvirziena nodošana formā plazmīda kontaktā baktēriju šūnas.
Plazmīdas ir mazas DNS molekulas. Tie nav saistīti ar hromosomu genomu un spēj autonomi dublēt. Plazmīdas satur gēnus, kas palielina baktēriju šūnu rezistenci pret nelabvēlīgiem vides apstākļiem. Baktērijas bieži nodod šos gēnus viena otrai. Tiek atzīmēta arī gēnu informācijas nodošana citas sugas baktērijām.
Ja nav patiesa seksuāla procesa, konjugācijai ir liela nozīme noderīgu īpašību apmaiņā. Tas pārnes baktēriju spēju izrādīt rezistenci pret zālēm. Cilvēcei īpaši bīstama ir antibiotiku rezistences pārnešana starp slimību izraisošām populācijām.

Rīsi. 17. Fotoattēlā divu Escherichia coli konjugācijas brīdis.
Baktēriju populācijas attīstības fāzes
Sējot uz barotnes, baktēriju populācijas attīstība notiek vairākos posmos.
Sākotnējā fāze
Sākotnējā fāze ir periods no sēšanas brīža līdz to augšanai. Vidēji sākuma fāze ilgst 1-2 stundas.
Reproduktīvās kavēšanās fāze
Šī ir baktēriju intensīvas augšanas fāze. Tās ilgums ir aptuveni 2 stundas. Tas ir atkarīgs no kultūras vecuma, adaptācijas perioda, barotnes kvalitātes utt.
logaritmiskā fāze
Šajā fāzē tiek atzīmēts reprodukcijas ātruma maksimums un baktēriju populācijas pieaugums. Tās ilgums ir 5-6 stundas.
Negatīvā paātrinājuma fāze
Šajā fāzē tiek novērota reprodukcijas ātruma samazināšanās, sadalošo baktēriju skaits samazinās un mirušo baktēriju skaits palielinās. Negatīvā paātrinājuma iemesls ir barības vielu barotnes izsīkums. Tās ilgums ir aptuveni 2 stundas.
Stacionāra maksimālā fāze
Stacionārajā fāzē tiek atzīmēts vienāds mirušo un jaunizveidoto indivīdu skaits. Tās ilgums ir aptuveni 2 stundas.
Paātrināta nāves fāze
Šajā posmā mirušo šūnu skaits pakāpeniski palielinās. Tās ilgums ir aptuveni 3 stundas.
Logaritmiskās nāves fāze
Šajā fāzē baktēriju šūnas mirst nemainīgā ātrumā. Tās ilgums ir aptuveni 5 stundas.
Samazināšanās fāze
Šajā fāzē atlikušās dzīvās baktēriju šūnas nonāk neaktīvā stāvoklī.

Rīsi. 18. Attēlā parādīta baktēriju populācijas augšanas līkne.

Rīsi. 19. Fotoattēlā redzamas zili zaļas Pseudomonas aeruginosa kolonijas, dzeltenās mikrokoku kolonijas, asinssarkanās Bacterium prodigiosum kolonijas un melnās Bacteroides niger kolonijas.

Rīsi. 20. Fotoattēlā redzama baktēriju kolonija. Katra kolonija ir vienas šūnas pēcnācējs. Kolonijā šūnu skaits ir miljons. kolonija izaug 1-3 dienās.
Magnētiski jutīgo baktēriju sadalīšana
1970. gados tika atklātas jūrās dzīvojošas baktērijas, kurām piemīt magnētisma sajūta. Magnētisms ļauj šīm apbrīnojamajām radībām pārvietoties pa Zemes magnētiskā lauka līnijām un atrast sēru, skābekli un citas tam tik nepieciešamās vielas. Viņu "kompasu" attēlo magnetosomas, kas sastāv no magnēta. Sadalot, magnētiski jutīgas baktērijas sadala savu kompasu. Šajā gadījumā sašaurināšanās dalīšanās laikā kļūst nepārprotami nepietiekama, tāpēc baktēriju šūna izliecas un rada asu lūzumu.

Rīsi. 21. Fotoattēlā redzams magnētiski jutīgas baktērijas dalīšanās brīdis.
Baktēriju augšana
Baktēriju šūnu dalīšanās sākumā divas DNS molekulas novirzās uz dažādiem šūnas galiem. Tālāk šūna tiek sadalīta divās vienādās daļās, kuras atdala viena no otras un palielina līdz sākotnējam izmēram. Daudzu baktēriju sadalīšanās ātrums ir vidēji 20-30 minūtes. Tikai vienas dienas laikā viena šūna veido 72 pēcnācēju paaudzes.
Šūnu masa augšanas un attīstības procesā ātri uzsūc barības vielas no vidi. To veicina labvēlīgi vides faktori – temperatūra, pietiekams barības vielu daudzums, nepieciešamais vides pH. Aerobām šūnām ir nepieciešams skābeklis. Anaerobiem tas ir bīstami. Taču neierobežota baktēriju vairošanās dabā nenotiek. Saules gaisma, sauss gaiss, barības trūkums, augsta apkārtējās vides temperatūra un citi faktori nelabvēlīgi ietekmē baktēriju šūnu.

Rīsi. 22. Fotogrāfijā šūnu dalīšanās brīdis.
augšanas faktori
Baktēriju augšanai nepieciešamas noteiktas vielas (augšanas faktori), no kurām daļu sintezē pati šūna, bet daļa nāk no apkārtējās vides. Visām baktērijām ir atšķirīgas prasības augšanas faktoriem.
Nepieciešamība pēc augšanas faktoriem ir nemainīga iezīme, kas ļauj to izmantot baktēriju identificēšanai, barotņu sagatavošanai un izmantošanai biotehnoloģijā.
Baktēriju augšanas faktori (bakteriālie vitamīni) ir ķīmiskie elementi, no kuriem lielākā daļa ir ūdenī šķīstošie B vitamīni.Šajā grupā ietilpst arī hemīna, holīna, purīna un pirimidīna bāzes, kā arī citas aminoskābes. Ja nav augšanas faktoru, notiek bakteriostāze.
Baktērijas izmanto augšanas faktorus minimālā daudzumā un nemainīgā veidā. Vairākas ķīmiskās vielas šajā grupā ir daļa no šūnu enzīmiem.

Rīsi. 23. Fotogrāfijā nūjiņveida baktērijas dalīšanās brīdis.
Svarīgākie baktēriju augšanas faktori
- B1 vitamīns (tiamīns). Piedalās ogļhidrātu metabolismā.
- B2 vitamīns (riboflavīns). Piedalās redoksreakcijās.
- Pantotēnskābe ir neatņemama koenzīma A sastāvdaļa.
- B6 vitamīns (piridoksīns). Piedalās aminoskābju metabolismā.
- Vitamīni B12(kobalamīni ir kobaltu saturošas vielas). Viņi aktīvi piedalās nukleotīdu sintēzē.
- Folijskābe. Daži no tā atvasinājumiem ir daļa no fermentiem, kas katalizē purīna un pirimidīna bāzes, kā arī dažu aminoskābju sintēzi.
- Biotīns. Piedalās slāpekļa metabolismā, kā arī katalizē nepiesātināto taukskābju sintēzi.
- PP vitamīns(nikotīnskābe). Piedalās redoksreakcijās, enzīmu veidošanā un lipīdu un ogļhidrātu metabolismā.
- H vitamīns(paraaminobenzoskābe). Tas ir augšanas faktors daudzām baktērijām, tostarp tām, kas mīt cilvēka zarnās. Folijskābe tiek sintezēta no para-aminobenzoskābes.
- Dvīņi. Tā ir dažu enzīmu, kas piedalās oksidācijas reakcijās, sastāvdaļa.
- Holīns. Piedalās šūnu sienas lipīdu sintēzes reakcijās. Tas ir metilgrupas piegādātājs aminoskābju sintēzē.
- Purīna un pirimidīna bāzes(adenīns, guanīns, ksantīns, hipoksantīns, citozīns, timīns un uracils). Vielas ir nepieciešamas galvenokārt kā nukleīnskābju sastāvdaļas.
- Aminoskābes. Šīs vielas ir šūnu proteīnu sastāvdaļas.
Nepieciešamība pēc dažu baktēriju augšanas faktoriem
Auksotrofi lai nodrošinātu dzīvību, viņiem nepieciešama ķīmisko vielu piegāde no ārpuses. Piemēram, klostridijas nespēj sintezēt lecitīnu un tirozīnu. Stafilokokiem ir nepieciešams uzņemt lecitīnu un arginīnu. Streptokokiem ir nepieciešams uzņemt taukskābes - fosfolipīdu sastāvdaļas. Korinebaktērijām un Shigella ir nepieciešama nikotīnskābes uzņemšana. Staphylococcus aureus, pneimokoku un brucelai nepieciešama B1 vitamīna uzņemšana. Streptokoki un stingumkrampju baciļi - pantotēnskābē.
Prototrofi patstāvīgi sintezēt nepieciešamās vielas.

Rīsi. 24. Dažādi apstākļi vides faktori dažādos veidos ietekmē baktēriju koloniju augšanu. Kreisajā pusē - stabila izaugsme lēni izplešanās apļa veidā. Labajā pusē - strauja izaugsme "dzinumu" veidā.
Baktēriju nepieciešamības pēc augšanas faktoriem izpēte ļauj zinātniekiem iegūt lielu mikrobu masu, kas ir tik nepieciešama pretmikrobu līdzekļu, serumu un vakcīnu ražošanā.
Vairāk par baktērijām lasiet rakstos:
Baktēriju vairošanās ir mehānisms mikrobu populāciju skaita palielināšanai. Baktēriju dalīšanās ir galvenais reprodukcijas veids. Pēc sadalīšanas baktērijām vajadzētu sasniegt pieaugušo izmēru. Baktērijas aug, ātri absorbējot barības vielas no savas vides. Augšanai nepieciešamas noteiktas vielas (augšanas faktori), no kurām daļu sintezē pati baktēriju šūna, bet daļa nāk no vides.
Pētot baktēriju augšanu un vairošanos, zinātnieki pastāvīgi atklāj labvēlīgās īpašības mikroorganismi, kuru izmantošana Ikdiena un ražošanā ierobežo tikai to īpašības.
Baktēriju darbību raksturo augšana- šūnas strukturālo un funkcionālo komponentu veidošanās un pašas baktēriju šūnas palielināšanās, kā arī vairošanās-- pašvairošanās, kas izraisa baktēriju šūnu skaita pieaugumu populācijā.
baktērijas vairojas ar bināro sadalīšanos uz pusēm, retāk ar pumpuru veidošanos. Aktinomicīti, tāpat kā sēnītes, var vairoties ar sporām. Aktinomicīti, kas ir zarojošas baktērijas, vairojas, sadrumstalojot pavedienveida šūnas. Grampozitīvās baktērijas sadalās, audzējot šūnā sintezētās dalīšanas starpsienas, un gramnegatīvās baktērijas sadalās ar sašaurināšanos, veidojoties hanteles formas figūrām, no kurām veidojas divas identiskas šūnas.
Pirms tam bija šūnu dalīšanās baktēriju hromosomas replikācija atbilstoši daļēji konservatīvam tipam (atveras divpavedienu DNS ķēde un katru virkni pabeidz komplementāra virkne), kas noved pie baktēriju kodola - nukleoīda - DNS molekulu dubultošanās.
DNS replikācija notiek trīs posmos: iniciācija, pagarināšana vai ķēdes augšana un izbeigšana.
Baktēriju pavairošana šķidrā barotnē. Baktērijas, kas iesētas noteiktā, nemainīgā barības barotnes tilpumā, vairojoties, patērē barības vielas, kas pēc tam noved pie barotnes izsīkuma un baktēriju augšanas pārtraukšanas. Baktēriju audzēšanu šādā sistēmā sauc par periodisku kultivēšanu, bet kultūru - par periodisku. Ja kultivēšanas apstākļi tiek uzturēti ar nepārtrauktu svaigas barotnes padevi un tāda paša tilpuma kultivēšanas šķidruma aizplūšanu, tad šādu kultivēšanu sauc par nepārtrauktu, un kultūru sauc par nepārtrauktu.
Audzējot baktērijas uz šķidras barotnes, tiek novērota gandrīz dibena, difūza vai virsmas (plēves veidā) kultūras augšana. Periodiskas baktēriju kultūras augšana, kas audzēta šķidrā barotnē, ir sadalīta vairākās fāzēs jeb periodos:
- 1. nobīdes fāze;
- 2. logaritmiskās augšanas fāze;
- 3. stacionāras augšanas fāze jeb maksimālā koncentrācija
baktērijas;
4. baktēriju nāves fāze.
Šīs fāzes var grafiski attēlot kā baktēriju reprodukcijas līknes segmentus, kas atspoguļo dzīvo šūnu skaita logaritma atkarību no to kultivēšanas laika.
Kavēšanās fāze- periods starp baktēriju sēšanu un vairošanās sākumu. Lagas fāzes ilgums ir vidēji 4-5 stundas.Baktērijas palielinās izmēros un gatavojas dalīšanai; palielinās nukleīnskābju, olbaltumvielu un citu komponentu daudzums.
Logaritmiskā (eksponenciālā) augšanas fāze ir intensīvas baktēriju dalīšanās periods. Tās ilgums ir aptuveni 5-6 stundas Optimālos augšanas apstākļos baktērijas var dalīties ik pēc 20-40 minūtēm. Šajā fāzē baktērijas ir visneaizsargātākās, kas izskaidrojams ar strauji augošas šūnas vielmaiņas komponentu augsto jutību pret proteīnu sintēzes inhibitoriem, nukleīnskābēm utt.
Tad nāk stacionāra augšanas fāze., kurā dzīvotspējīgo šūnu skaits paliek nemainīgs, veidojot maksimālo līmeni (M-koncentrāciju). Tās ilgums ir izteikts stundās un mainās atkarībā no baktēriju veida, to īpašībām un audzēšanas.
Nāves fāze pabeidz baktēriju augšanas procesu, ko raksturo baktēriju nāve uzturvielu barotnes avotu izsīkuma apstākļos un baktēriju vielmaiņas produktu uzkrāšanās tajā. Tās ilgums svārstās no 10 stundām līdz vairākām nedēļām. Baktēriju augšanas un vairošanās intensitāte ir atkarīga no daudziem faktoriem, tostarp no optimālā uzturvielu barotnes sastāva, redokspotenciāla, pH, temperatūras utt.
Baktēriju pavairošana blīvā barotnē. Baktērijas, kas aug uz blīvām barotnēm, veido izolētas noapaļotas kolonijas ar vienmērīgām vai nelīdzenām malām (S- un R-formas), ar dažādu konsistenci un krāsu atkarībā no baktēriju pigmenta.
Ūdenī šķīstošie pigmenti izkliedējas uzturvielu vidē un krāso to. Vēl viena pigmentu grupa nešķīst ūdenī, bet šķīst organiskajos šķīdinātājos. Un, visbeidzot, ir pigmenti, kas nešķīst ne ūdenī, ne organiskajos savienojumos.
Visizplatītākie mikroorganismu pigmenti ir karotīni, ksantofili un melanīni. Melanīni ir nešķīstoši melni, brūni vai sarkani pigmenti, kas sintezēti no fenola savienojumiem. Melanīni kopā ar katalāzi, superoksīda cismutāzi un peroksidāzēm aizsargā mikroorganismus no toksisko skābekļa peroksīda radikāļu iedarbības. Daudziem pigmentiem ir pretmikrobu, antibiotikām līdzīga iedarbība.