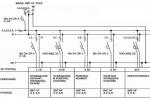
Baktēriju šūnu struktūra
Baktēriju struktūra ir labi pētīta, izmantojot elektronu mikroskopiju veselām šūnām un to īpaši plānām sekcijām. Baktēriju šūna sastāv no šūnas sienas, citoplazmas membrānas, citoplazmas ar ieslēgumiem un kodola, ko sauc par nukleoīdu. Ir papildu struktūras: kapsula, mikrokapsula, gļotas, flagellas, pili (1. att.); Dažas baktērijas nelabvēlīgos apstākļos spēj veidot sporas.
Šūnapvalki - spēcīga, elastīga struktūra, kas piešķir baktērijai noteiktu formu un kopā ar pamatā esošo citoplazmas membrānu “ierobežo” augsto osmotisko spiedienu baktēriju šūna. Tas ir iesaistīts šūnu dalīšanās procesā un metabolītu transportēšanā. Visbiezākā šūnu siena ir grampozitīvām baktērijām (1. att.). Tātad, ja gramnegatīvo baktēriju šūnu sienas biezums ir aptuveni 15-20 nm, tad grampozitīvās baktērijās tas var sasniegt 50 nm vai vairāk. Grampozitīvo baktēriju šūnu siena satur nelielu daudzumu polisaharīdu, lipīdu un olbaltumvielu.
Šo baktēriju šūnu sienas galvenā sastāvdaļa ir daudzslāņu peptidoglikāns(mureīns, mukopeptīds), kas veido 40-90% no šūnas sienas masas.
Volutina mezosomas nukleoīds
Rīsi. 1. Baktēriju šūnas uzbūve.
Teihoīnskābes (no grieķu valodas. teichos - siena), kuru molekulas ir 8-50 glicerīna un ribitola atlieku ķēdes, kas savienotas ar fosfātu tiltiem. Baktēriju formu un stiprumu nosaka stingrā peptidoglikāna šķiedru struktūra, kas ir daudzslāņu un šķērssavienota ar peptīdiem. Peptidoglikānu attēlo paralēlas glikāna molekulas, kas sastāv no atkārtotiem atlikumiem N-acetilglikozamīns un N-acetilmuramīnskābe, kas savienota ar P tipa glikozīdu saiti (1 -> 4).
Lizocīms, kas ir acetilmuramidāze, sarauj šīs saites. Glikāna molekulas ir saistītas ar peptīdu šķērssavienojumu. Līdz ar to šī polimēra nosaukums - peptidoglikāns. Peptidoglikāna peptīdu saites pamatā gramnegatīvās baktērijās ir tetrapeptīdi, kas sastāv no mainīgiem L- Un D-aminoskābes.
U E. coli peptīdu ķēdes ir savienotas viena ar otru caur D- vienas ķēdes alanīns un otras mezodiaminopimelskābe.
Peptidoglikāna peptīdu daļas sastāvs un struktūra gramnegatīvajās baktērijās ir stabila, atšķirībā no grampozitīvo baktēriju peptidoglikāna, kuru aminoskābes var atšķirties pēc sastāva un secības. Tetrapeptīdi šeit ir savienoti viens ar otru ar 5 glicīna atlikumu polipeptīdu ķēdēm. Grampozitīvās baktērijas bieži satur lizīnu, nevis mezodiaminopimelskābi. Fosfolipīds

Rīsi. 2. Grampozitīvo (gram+) un gramnegatīvo (gram") baktēriju virsmas struktūru struktūra.
Glikāna elementi (acetilglikozamīns un acetilmuramīnskābe) un tetrapeptīdu aminoskābes (mezodiaminopimēlskābe un L-glutamīnskābe, D-alanīns) ir baktēriju atšķirīga iezīme, jo dzīvniekiem un cilvēkiem tie un aminoskābju D-izomēri nav sastopami.
Grampozitīvo baktēriju spēja saglabāt genciānas violetu kombinācijā ar jodu, krāsojot, izmantojot Grama traipu (zili violeta baktēriju krāsa), ir saistīta ar daudzslāņu peptidoglikāna īpašību mijiedarboties ar krāsvielu. Turklāt turpmāka baktēriju uztriepes apstrāde ar spirtu izraisa peptidoglikāna poru sašaurināšanos un tādējādi krāsvielas aizturi šūnu sieniņā. Pēc alkohola iedarbības gramnegatīvās baktērijas zaudē savu krāsvielu, maina krāsu un, apstrādājot ar fuksīnu, kļūst sarkanas. Tas ir saistīts ar mazāku peptidoglikāna daudzumu (5-10% no šūnu sienas masas).
Gramnegatīvo baktēriju šūnu siena satur ārējā membrāna, caur lipoproteīnu savienots ar apakšējo peptidoglikāna slāni (2. att.). Ārējā membrāna ir viļņota trīsslāņu struktūra, līdzīga iekšējai membrānai, ko sauc par citoplazmatisku. Šo membrānu galvenā sastāvdaļa ir bimolekulārais (dubultais) lipīdu slānis.
Ārējā membrāna ir asimetriska mozaīkas struktūra, ko attēlo lipopolisaharīdi, fosfolipīdi un olbaltumvielas . Tās ārējā pusē ir lipopolisaharīds(LPS), kas sastāv no trim sastāvdaļām: lipīdu A, serdes daļa vai serde (lat. kodols - kodols) un 0-specifiska polisaharīdu ķēde, kas veidojas, atkārtojot oligosaharīdu sekvences.
Lipopolisaharīds ir “noenkurots” ārējā membrānā ar lipīdu palīdzību A, izraisot LPS toksicitāti, kas tādējādi tiek identificēta ar endotoksīnu. Baktēriju iznīcināšana ar antibiotikām izraisa lielu endotoksīna daudzumu izdalīšanos, kas pacientam var izraisīt endotoksisku šoku.
No lipīdiem A kodols vai LPS galvenā daļa atdalās. LPS kodola konstantākā daļa ir ketodeoksioktonskābe (3-deoksi-g)-manno-2-oktulozonskābe). 0 -nosaka specifiska ķēde, kas stiepjas no LPS molekulas kodola daļas serogrupa, serovars (baktēriju veids, kas noteikts, izmantojot imūnserumu) specifisks baktēriju celms. Tādējādi LPS jēdziens ir saistīts ar 0-antigēna jēdzienu, ko var izmantot baktēriju diferencēšanai. Ģenētiskās izmaiņas var izraisīt izmaiņas komponentu biosintēzē LPS baktērijas un rezultātā L- veidlapas
Matricas proteīniārējā membrāna iekļūst tajā tādā veidā, ka olbaltumvielu molekulas sauc porinami, robežojas ar hidrofilām porām, caur kurām iziet ūdens un mazas molekulas ar relatīvo masu līdz 700. Starp ārējo un citoplazmas membrānu atrodas periplazmatiska telpa jeb periplazma, kas satur fermentus. Ja lizocīma, penicilīna, ķermeņa aizsargfaktoru un citu savienojumu ietekmē tiek traucēta baktēriju šūnu sienas sintēze, veidojas šūnas ar mainītu (bieži sfērisku) formu: protoplasti - baktērijas, kurām pilnībā trūkst šūnu sienas; sferoplasti - baktērijas ar daļēji saglabātu šūnu sieniņu. Pēc šūnu sienas inhibitora noņemšanas šādas izmainītas baktērijas var mainīties, t.i. iegūst pilnu šūnu sieniņu un atjauno tās sākotnējo formu.
Sfero- vai protoplasta tipa baktērijas, kuras antibiotiku vai citu faktoru ietekmē ir zaudējušas spēju sintezēt peptidoglikānu un ir spējīgas vairoties, sauc. L formas(no Listera institūta nosaukuma). L-formas var rasties arī mutāciju rezultātā. Tās ir osmotiski jutīgas, sfēriskas, dažāda izmēra kolbas formas šūnas, arī tās, kas iziet cauri baktēriju filtriem. Dažas L- formas (nestabils), kad tiek noņemts faktors, kas izraisīja izmaiņas baktērijās, var mainīties, “atgriežoties” sākotnējā baktēriju šūnā. L- formas var veidot daudzi infekcijas slimību patogēni.
Citoplazmas membrāna ultraplānu sekciju elektronmikroskopijā tā ir trīsslāņu membrāna, kas ieskauj baktēriju citoplazmas ārējo daļu. Pēc struktūras tas ir līdzīgs dzīvnieku šūnu plazmalemmai un sastāv no dubultā lipīdu slāņa, galvenokārt fosfolipīdiem ar iestrādātu virsmu un integrāliem proteīniem, kas, šķiet, iekļūst caur membrānas struktūru. Dažas no tām ir caurlaides, kas iesaistītas vielu transportēšanā. Citoplazmas membrāna ir dinamiska struktūra ar mobilām sastāvdaļām, tāpēc to uzskata par mobilu šķidruma struktūru. Tas ir iesaistīts osmotiskā spiediena regulēšanā, vielu transportēšanā un šūnas enerģijas metabolismā (sakarā ar elektronu transportēšanas ķēdes enzīmiem, adenozīna trifosfatāzi utt.). Ar pārmērīgu augšanu (salīdzinājumā ar šūnu sienas augšanu) citoplazmas membrāna veido invaginācijas - invaginācijas sarežģīti savītu membrānu struktūru veidā, t.s. mezosomas. Tiek sauktas mazāk sarežģītas savītas struktūras intracitoplazmas membrānas. Mezosomu un intracitoplazmas membrānu loma nav pilnībā izprotama. Tiek pat ierosināts, ka tie ir artefakts, kas rodas pēc parauga sagatavošanas (fiksēšanas) elektronu mikroskopijai. Neskatoties uz to, tiek uzskatīts, ka citoplazmas membrānas atvasinājumi piedalās šūnu dalīšanās procesā, nodrošinot enerģiju šūnas sieniņas sintēzei, un piedalās vielu sekrēcijā, sporulācijā, t.i. procesos ar augstu enerģijas patēriņu.
Citoplazma aizņem lielāko daļu baktēriju šūnas un sastāv no šķīstošiem proteīniem, ribonukleīnskābēm, ieslēgumiem un daudzām mazām granulām - ribosomas atbildīgs par olbaltumvielu sintēzi (tulkošanu). Baktēriju ribosomu izmērs ir aptuveni 20 nm un sedimentācijas koeficients 70S, 3 atšķirība no 80^-ribosomām, kas raksturīgas eikariotu šūnām. Tāpēc dažas antibiotikas, saistoties ar baktēriju ribosomām, nomāc baktēriju proteīnu sintēzi, neietekmējot proteīnu sintēzi eikariotu šūnās. Baktēriju ribosomas var sadalīties divās apakšvienībās - 50S Un 30S . Citoplazmā ir dažādi ieslēgumi glikogēna granulu, polisaharīdu, poli-p-sviestskābes un polifosfātu (volutīna) veidā. Tie uzkrājas, kad vidē ir barības vielu pārpalikums, un darbojas kā rezerves vielas uzturam un enerģijas vajadzībām. Volutin ir afinitāte pret pamata krāsvielām, tai ir metahromāzija, un to ir viegli noteikt, izmantojot īpašas krāsošanas metodes. Volutīna graudu raksturīgais izvietojums atklājas difterijas nūjiņā intensīvi iekrāsotu šūnu polu veidā.
Nukleoīds - līdzvērtīgs baktēriju kodolam. Tas atrodas baktēriju centrālajā zonā divpavedienu DNS veidā, noslēgts gredzenā un cieši saspiests kā bumba. Atšķirībā no eikariotiem, baktēriju kodolam nav kodola apvalka, kodola vai pamata proteīnu (histonu). Parasti baktēriju šūnā ir viena hromosoma, ko attēlo DNS molekula, kas noslēgta gredzenā. Ja dalīšanās ir traucēta, tajā var būt 4 vai vairāk hromosomas. Nukleoīdu nosaka gaismas mikroskopā pēc krāsošanas, izmantojot DNS specifiskās metodes: Feulgen vai Romanovsky-Giemsa. Ultraplānas baktēriju sekciju elektronu difrakcijas modeļos nukleoīds parādās kā gaismas zonas ar fibrilārām, pavedieniem līdzīgām DNS struktūrām, kas noteiktos apgabalos ir saistītas ar citoplazmas membrānu vai mezosomu, kas iesaistīta hromosomu replikācijā.
Papildus nukleoīdam, ko pārstāv viena hromosoma, baktēriju šūna satur ekstrahromosomu iedzimtības faktorus - plazmīdas, kas ir kovalenti noslēgti DNS gredzeni.
Kapsula - gļotādas struktūra, kas biezāka par 0,2 mikroniem, cieši saistīta ar baktēriju šūnu sieniņu un ar skaidri noteiktām ārējām robežām. Kapsula ir redzama nospieduma uztriepes no patoloģiskā materiāla. Tīrās baktēriju kultūrās kapsula veidojas retāk. To nosaka, izmantojot īpašas Burri-Gins krāsošanas metodes, kas rada kapsulas vielu negatīvu kontrastu.
Parasti kapsula sastāv no polisaharīdiem (eksopolisaharīdiem), dažreiz no polipeptīdiem, piemēram, Sibīrijas mēra nūjiņā. Kapsula ir hidrofila, tā novērš baktēriju fagocitozi.
Veidojas daudzas baktērijas mikrokapsula - gļotādas veidošanās, kuras biezums ir mazāks par 0,2 mikroniem, nosakāms tikai ar elektronu mikroskopu. To vajadzētu atšķirt no kapsulas gļotas - gļotādas eksopolisaharīdi, kuriem nav skaidras ārējās robežas. Gļotādas eksopolisaharīdi ir raksturīgi Pseudomonas aeruginosa mukoīdiem celmiem, kas bieži sastopami cistiskās fibrozes pacientu krēpās. Baktēriju eksopolisaharīdi ir iesaistīti adhēzijā (pielīp pie substrātiem); tos sauc arī par glikokalikss. Papildus baktēriju veiktajai eksopolisaharīdu sintēzei ir vēl viens to veidošanās mehānisms: ekstracelulāro baktēriju enzīmu iedarbība uz disaharīdiem. Tā rezultātā veidojas dekstrāni un levāni. Kapsula un gļotas aizsargā baktērijas no bojājumiem un izžūšanas, jo, būdami hidrofilas, tās labi saista ūdeni un novērš makroorganisma un bakteriofāgu aizsargfaktoru darbību.
Flagella baktērijas nosaka baktēriju šūnas mobilitāti. Flagellas ir plāni pavedieni, kas nāk no citoplazmas membrānas un ir garāki par pašu šūnu (3. att.). Ziedu biezums ir 12-20 nm, garums 3-12 µm. Ziedu skaits dažādās baktēriju sugās atšķiras no viena (monotrich) holera vibrio ir līdz pat desmitiem un simtiem flagellas, kas stiepjas gar baktērijas perimetru (peri-trich) Escherichia coli, Proteus u.c. Lofotrihs vienā šūnas galā ir karogs saišķis. Amfitrihijašūnas pretējos galos ir viens flagellum vai ziepju saišķis. Flagellas ir piestiprinātas pie citoplazmas membrānas un šūnu sienas ar īpašiem diskiem. Flagella sastāv no proteīna - flagellīna (no naT.flagellum — flagellum), kam piemīt antigēnu specifiskums. Flagellina apakšvienības ir savītas spirāles formā. Ziedi tiek atklāti, izmantojot elektronu mikroskopiju preparātiem, kas pārklāti ar smagajiem metāliem, vai gaismas mikroskopā pēc apstrādes ar īpašām metodēm, kuru pamatā ir dažādu vielu kodināšana un adsorbcija, kas izraisa flagellas biezuma palielināšanos (piemēram, pēc sudraba).

Rīsi. 3. Escherichia coli. Elektronu difrakcijas modelis (sagatavots V.S. Tyurin). 1 - flagella, 2 - villi, 3 - F-pili.
Villi jeb pili (fimbriae), - pavedieniem līdzīgi veidojumi (3. att.), tievāki un īsāki (3-10 nm x 0,3-10 µm) nekā flagellas. Pili stiepjas no šūnas virsmas un sastāv no proteīna pilīna. Viņiem ir antigēna aktivitāte. Starp pili ir: pili, kas atbild par adhēziju, t.i. baktēriju pievienošanai skartajai šūnai (1. tipa pili vai vispārējais tips - kopējā pili), dzēra, atbild par uzturu, ūdens-sāls metabolismu; seksuāla (F-dzēra), vai konjugācijas pili (2. tipa pili). Vispārējā tipa pili ir daudz - vairāki simti katrā šūnā. Dzimuma pili veido tā sauktās “vīriešu” donoru šūnas, kas satur transmisīvās plazmīdas (F, R, Col). Parasti vienā šūnā tās ir 1-3. Dzimuma pili atšķirīga iezīme ir mijiedarbība ar īpašiem “vīriešu” sfēriskiem bakteriofāgiem, kas intensīvi adsorbējas uz dzimuma pilieniem.
Pretrunas - savdabīga miera stāvoklī esošo firmicute baktēriju forma, t.i. baktērijas ar grampozitīvu šūnu sienas struktūru.
Sporas veidojas baktēriju pastāvēšanai nelabvēlīgos apstākļos (žāvēšana, barības vielu deficīts utt.). Šajā gadījumā vienas baktērijas iekšpusē veidojas viena spora. Sporu veidošanās veicina sugas saglabāšanos un nav vairošanās metode, kā tas ir sēnēs.
Dažkārt tiek sauktas sporas veidojošas aerobās baktērijas, kurās sporu izmērs nepārsniedz šūnas diametru baciļi. Tiek sauktas sporas veidojošās anaerobās baktērijas, kurās sporu izmērs pārsniedz šūnas diametru un tāpēc iegūst vārpstas formu. klostridijas(lat. klostridijs- vārpsta).
Process sporulācija(sporulācija) iziet vairākus posmus, kuru laikā daļa citoplazmas un hromosoma tiek atdalīta, ko ieskauj citoplazmas membrāna; Veidojas prospora, tad veidojas daudzslāņu, slikti caurlaidīgs apvalks. Sporulāciju pavada intensīvs prosporu patēriņš un pēc tam dipikolīnskābes un kalcija jonu sporu apvalka veidošanās. Pēc visu struktūru veidošanās sporas iegūst karstumizturību, kas ir saistīta ar kalcija dipikolināta klātbūtni. Sporulācija, sporu forma un atrašanās vieta šūnā (veģetatīvā) ir baktēriju sugas īpašība, kas ļauj tās atšķirt vienu no otras. Sporu forma var būt ovāla, sfēriska, atrašanās šūnā ir termināla, t.i. nūjas galā (stingumkrampju izraisītājs), apakšgalā - tuvāk nūjas galam (botulisma patogēni, gāzes gangrēna) un centrālajā (sibīrijas mēra bacilis).
Baktērijas ir vecākā organismu grupa, kas pašlaik pastāv uz Zemes. Pirmās baktērijas, iespējams, parādījās pirms vairāk nekā 3,5 miljardiem gadu, un gandrīz miljardu gadu tās bija vienīgās dzīvās būtnes uz mūsu planētas. Tā kā šie bija pirmie dzīvās dabas pārstāvji, viņu ķermenim bija primitīva uzbūve.
Laika gaitā to struktūra kļuva sarežģītāka, taču līdz pat šai dienai baktērijas tiek uzskatītas par primitīvākajiem vienšūnas organismiem. Interesanti, ka dažas baktērijas joprojām saglabā savu seno senču primitīvās iezīmes. To novēro baktērijās, kas dzīvo karstos sēravotos un bezskābekļa dubļos rezervuāru apakšā.
Lielākā daļa baktēriju ir bezkrāsainas. Tikai daži ir violeti vai zaļi. Bet daudzu baktēriju kolonijām ir spilgta krāsa, ko izraisa krāsainas vielas izdalīšanās vidē vai šūnu pigmentācija.
Baktēriju pasaules atklājējs bija 17. gadsimta holandiešu dabaszinātnieks Antonijs Lēvenhuks, kurš pirmais radīja perfektu palielināmo mikroskopu, kas objektus palielina 160-270 reizes.
Baktērijas tiek klasificētas kā prokariotes un tiek klasificētas atsevišķā valstībā - Baktērijas.
Ķermeņa forma
Baktērijas ir daudz un dažādi organismi. Tās atšķiras pēc formas.
| Baktērijas nosaukums | Baktēriju forma | Baktēriju attēls |
| Cocci | Lodveida | |
| Bacillus |  | Stieņa formas |
| Vibrio | Komatveida | |
| Spirillum |  | Spirāle |
| Streptokoki |  | Koku ķēde |
| Stafilokoks |  | Koku kopas |
| Diplokoks | Divas apaļas baktērijas, kas ievietotas vienā gļotādas kapsulā |
Pārvadāšanas metodes
Starp baktērijām ir mobilās un nekustīgās formas. Kustības pārvietojas viļņveidīgu kontrakciju dēļ vai ar flagellas (savītu spirālveida pavedienu) palīdzību, kas sastāv no īpaša proteīna, ko sauc par flagellīnu. Var būt viena vai vairākas flagellas. Dažās baktērijās tie atrodas vienā šūnas galā, citās - divos vai pa visu virsmu.

Bet kustība ir raksturīga arī daudzām citām baktērijām, kurām trūkst flagellas. Tādējādi baktērijas, kas no ārpuses pārklātas ar gļotām, spēj slīdēt.
Dažām ūdens un augsnes baktērijām, kurām nav flagellas, citoplazmā ir gāzes vakuoli. Šūnā var būt 40-60 vakuolu. Katrs no tiem ir piepildīts ar gāzi (domājams, slāpekli). Regulējot gāzes daudzumu vakuolos, ūdens baktērijas var iegrimt ūdens stabā vai pacelties uz tās virsmu, un augsnes baktērijas var pārvietoties augsnes kapilāros.
Dzīvotne
Savas organizācijas vienkāršības un nepretenciozitātes dēļ baktērijas ir plaši izplatītas dabā. Baktērijas ir sastopamas visur: pat vistīrākā avota ūdens pilē, augsnes graudos, gaisā, akmeņos, polārajā sniegā, tuksneša smiltīs, okeāna dibenā, eļļā, kas iegūta no liela dziļuma, un pat karsto avotu ūdens, kura temperatūra ir aptuveni 80ºC. Viņi dzīvo uz augiem, augļiem, dažādiem dzīvniekiem un cilvēka zarnās, mutes dobums, uz ekstremitātēm, uz ķermeņa virsmas.
Baktērijas ir mazākās un daudzskaitlīgākās dzīvās būtnes. Mazā izmēra dēļ tie viegli iekļūst plaisās, spraugās vai porās. Ļoti izturīgs un pielāgots dažādiem dzīves apstākļiem. Tie panes žāvēšanu, lielu aukstumu un karsēšanu līdz 90ºC, nezaudējot savu dzīvotspēju.
Uz Zemes praktiski nav tādas vietas, kur baktērijas netiktu atrastas, bet dažādos daudzumos. Baktēriju dzīves apstākļi ir dažādi. Dažiem no tiem nepieciešams atmosfēras skābeklis, citiem tas nav vajadzīgs un spēj dzīvot bezskābekļa vidē.
Gaisā: baktērijas paceļas augšējos atmosfēras slāņos līdz 30 km. un vēl.
Īpaši daudz to ir augsnē. 1 g augsnes var saturēt simtiem miljonu baktēriju.
Ūdenī: ūdens virszemes slāņos atklātos rezervuāros. Noderīgās ūdens baktērijas mineralizē organiskās atliekas.
Dzīvos organismos: patogēnās baktērijas nokļūst organismā no ārējās vides, bet tikai labvēlīgos apstākļos izraisa slimības. Simbiotiķi dzīvo gremošanas orgānos, palīdzot sadalīt un uzņemt pārtiku, sintezēt vitamīnus.
Ārējā struktūra
Baktērijas šūna ir pārklāta ar īpašu blīvu apvalku - šūnas sieniņu, kas veic aizsargfunkcijas un atbalsta funkcijas, kā arī piešķir baktērijai paliekošu, raksturīgu formu. Baktērijas šūnu siena atgādina augu šūnas sienu. Tas ir caurlaidīgs: caur to barības vielas brīvi nokļūst šūnā, un vielmaiņas produkti iziet vidē. Nereti baktērijas rada papildu aizsargslāni ar gļotām uz šūnas sieniņas – kapsulu. Kapsulas biezums var būt daudzkārt lielāks par pašas šūnas diametru, taču tas var būt arī ļoti mazs. Kapsula nav būtiska šūnas sastāvdaļa, tā veidojas atkarībā no apstākļiem, kādos atrodas baktērijas. Tas pasargā baktērijas no izžūšanas.
Dažu baktēriju virspusē ir garas karogs (viena, divas vai daudzas) vai īsas plānas bārkstiņas. Ziedu garums var būt daudzkārt lielāks par baktērijas ķermeņa izmēru. Baktērijas pārvietojas ar flagellas un bārkstiņu palīdzību.

Iekšējā struktūra
Baktēriju šūnas iekšpusē ir blīva, nekustīga citoplazma. Tam ir slāņaina struktūra, tajā nav vakuolu, tāpēc dažādas olbaltumvielas (enzīmi) un rezerves barības vielas atrodas pašā citoplazmas vielā. Baktēriju šūnām nav kodola. Viela, kas satur iedzimtu informāciju, ir koncentrēta viņu šūnas centrālajā daļā. Baktērijas, - nukleīnskābe - DNS. Bet šī viela nav izveidota kodolā.
Baktēriju šūnas iekšējā organizācija ir sarežģīta, un tai ir savas specifiskās īpašības. Citoplazmu no šūnas sienas atdala citoplazmas membrāna. Citoplazmā ir galvenā viela jeb matrica, ribosomas un neliels skaits membrānu struktūru, kas veic dažādas funkcijas (mitohondriju analogi, endoplazmatiskais tīkls, Golgi aparāts). Baktēriju šūnu citoplazmā bieži ir dažādu formu un izmēru granulas. Granulas var sastāvēt no savienojumiem, kas kalpo kā enerģijas un oglekļa avots. Tauku pilieni ir atrodami arī baktēriju šūnā.
Šūnas centrālajā daļā ir lokalizēta kodolviela - DNS, kas nav norobežota no citoplazmas ar membrānu. Tas ir kodola analogs - nukleoīds. Nukleoīdam nav membrānas, kodola vai hromosomu kopas.
Ēšanas metodes
Baktērijām ir dažādas barošanas metodes. Starp tiem ir autotrofi un heterotrofi. Autotrofi ir organismi, kas spēj patstāvīgi ražot organiskas vielas savai uzturam.
Augiem ir nepieciešams slāpeklis, bet paši nevar absorbēt slāpekli no gaisa. Dažas baktērijas apvieno gaisā esošās slāpekļa molekulas ar citām molekulām, kā rezultātā veidojas augiem pieejamas vielas.
Šīs baktērijas apmetas jauno sakņu šūnās, kā rezultātā uz saknēm veidojas sabiezējumi, ko sauc par mezgliņiem. Šādi mezgliņi veidojas uz pākšaugu dzimtas augu un dažu citu augu saknēm.
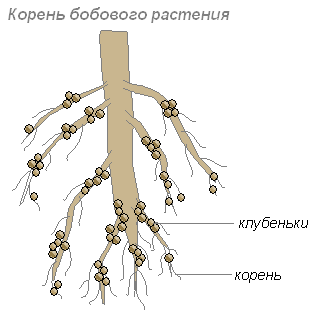
Saknes nodrošina baktērijām ogļhidrātus, bet baktērijas saknēm nodrošina slāpekli saturošas vielas, kuras augs var absorbēt. Viņu kopdzīve ir abpusēji izdevīga.
Augu saknes izdala daudz organisko vielu(cukuri, aminoskābes un citi), ar kuriem barojas baktērijas. Tāpēc īpaši daudz baktēriju apmetas augsnes slānī, kas ieskauj saknes. Šīs baktērijas pārvērš mirušos augu atliekas augiem pieejamās vielās. Šo augsnes slāni sauc par rizosfēru.
Pastāv vairākas hipotēzes par mezgliņu baktēriju iekļūšanu sakņu audos:
- caur epidermas un garozas audu bojājumiem;
- caur sakņu matiņiem;
- tikai caur jauno šūnu membrānu;
- pateicoties pavadošajām baktērijām, kas ražo pektinolītiskos enzīmus;
- B-indoletiķskābes sintēzes stimulēšanas dēļ no triptofāna, kas vienmēr atrodas augu sakņu sekrēcijās.
Mezglu baktēriju ievadīšanas process sakņu audos sastāv no divām fāzēm:
- sakņu matiņu infekcija;
- mezgliņu veidošanās process.
Vairumā gadījumu invāzijas šūna aktīvi vairojas, veido tā sauktos infekcijas pavedienus un šādu pavedienu veidā pārvietojas augu audos. Mezglu baktērijas, kas rodas no infekcijas pavediena, turpina vairoties saimniekaudi.

Augu šūnas, kas piepildītas ar ātri vairojošām mezgliņu baktēriju šūnām, sāk strauji dalīties. Jauna mezgliņa savienojums ar pākšaugu sakni tiek veikts, pateicoties asinsvadu-šķiedru saišķiem. Funkcionēšanas periodā mezgliņi parasti ir blīvi. Līdz brīdim, kad notiek optimāla aktivitāte, mezgliņi iegūst rozā krāsu (pateicoties leghemoglobīna pigmentam). Tikai tās baktērijas, kas satur leghemoglobīnu, spēj piesaistīt slāpekli.
Mezglu baktērijas rada desmitiem un simtiem kilogramu slāpekļa mēslojuma uz hektāru augsnes.
Vielmaiņa
Baktērijas atšķiras viena no otras savā vielmaiņā. Dažos tas notiek ar skābekļa piedalīšanos, citās - bez tā.
Lielākā daļa baktēriju barojas ar gatavām organiskām vielām. Tikai dažas no tām (zili zaļas vai zilaļģes) spēj radīt organiskas vielas no neorganiskām. Viņiem bija svarīga loma skābekļa uzkrāšanā Zemes atmosfērā.
Baktērijas absorbē vielas no ārpuses, saplēš to molekulas gabalos, no šīm daļām saliek čaulu un papildina to saturu (tā tās aug), un izmet nevajadzīgās molekulas. Baktērijas apvalks un membrāna ļauj tai tikai absorbēt nepieciešamās vielas.

Ja baktērijas apvalks un membrāna būtu pilnībā necaurlaidīgi, šūnā neiekļūtu nekādas vielas. Ja tie būtu caurlaidīgi visām vielām, šūnas saturs sajauktos ar barotni – šķīdumu, kurā dzīvo baktērija. Lai izdzīvotu, baktērijām ir nepieciešams apvalks, kas ļauj iziet cauri nepieciešamām vielām, bet ne nevajadzīgām vielām.
Baktērija absorbē barības vielas, kas atrodas tās tuvumā. Kas notiek tālāk? Ja tas var pārvietoties patstāvīgi (pārvietojot zizli vai atgrūžot gļotas), tad tas kustas, līdz atrod nepieciešamās vielas.

Ja tas nevar kustēties, tad gaida, kamēr difūzija (vienas vielas molekulu spēja iekļūt citas vielas molekulu biezoknī) atnes tai nepieciešamās molekulas.
Baktērijas kopā ar citām mikroorganismu grupām veic milzīgu ķīmisko darbu. Pārvēršot dažādus savienojumus, tie saņem dzīvībai nepieciešamo enerģiju un uzturvielas. Metabolisma procesi, enerģijas iegūšanas metodes un nepieciešamība pēc materiāliem to ķermeņa vielu veidošanai baktērijās ir dažādi.
Citas baktērijas apmierina visas savas vajadzības pēc oglekļa, kas nepieciešamas organisko vielu sintēzei organismā uz neorganisko savienojumu rēķina. Tos sauc par autotrofiem. Autotrofās baktērijas spēj sintezēt organiskās vielas no neorganiskām. Starp tiem ir:
Ķīmijsintēze
Starojuma enerģijas izmantošana ir vissvarīgākais, bet ne vienīgais veids, kā radīt organiskās vielas no oglekļa dioksīda un ūdens. Ir zināmas baktērijas, kas kā enerģijas avots šādai sintēzei izmanto enerģiju, nevis saules gaismu. ķīmiskās saites, kas rodas organismu šūnās noteiktu neorganisko savienojumu - sērūdeņraža, sēra, amonjaka, ūdeņraža, slāpekļskābes, dzelzs savienojumu un mangāna - oksidēšanās laikā. Viņi izmanto organiskās vielas, kas veidojas, izmantojot šo ķīmisko enerģiju, lai izveidotu sava ķermeņa šūnas. Tāpēc šo procesu sauc par ķīmijsintēzi.
Vissvarīgākā ķīmiski sintētisko mikroorganismu grupa ir nitrificējošās baktērijas. Šīs baktērijas dzīvo augsnē un oksidē amonjaku, kas veidojas organisko atlieku sadalīšanās laikā līdz slāpekļskābei. Pēdējais reaģē ar augsnes minerālu savienojumiem, pārvēršoties slāpekļskābes sāļos. Šis process notiek divos posmos.
Dzelzs baktērijas pārvērš dzelzi par dzelzi oksīdu. Iegūtais dzelzs hidroksīds nosēžas un veido tā saukto purva dzelzsrūdu.
Daži mikroorganismi pastāv molekulārā ūdeņraža oksidācijas dēļ, tādējādi nodrošinot autotrofisku uztura metodi.
Ūdeņraža baktēriju raksturīga iezīme ir spēja pāriet uz heterotrofisku dzīvesveidu, ja tās tiek nodrošinātas ar organiskiem savienojumiem un ūdeņraža neesamību.
Tādējādi ķīmijautotrofi ir tipiski autotrofi, jo tie patstāvīgi sintezē nepieciešamos organiskos savienojumus no neorganiskām vielām un neņem tos gatavus no citiem organismiem, piemēram, heterotrofiem. Ķīmijautotrofās baktērijas atšķiras no fototrofiskajiem augiem ar to pilnīgu neatkarību no gaismas kā enerģijas avota.
Baktēriju fotosintēze
Dažas pigmentu saturošas sēra baktērijas (violeta, zaļa), kas satur specifiskus pigmentus - bakteriohlorofilus, spēj absorbēt saules enerģija, ar kuras palīdzību sērūdeņradis to ķermeņos tiek sadalīts un atbrīvo ūdeņraža atomus, lai atjaunotu atbilstošos savienojumus. Šim procesam ir daudz kopīga ar fotosintēzi un tas atšķiras tikai ar to, ka purpursarkanajās un zaļajās baktērijās ūdeņraža donors ir sērūdeņradis (reizēm karbonskābes), bet zaļajos augos tas ir ūdens. Abos no tiem ūdeņraža atdalīšana un pārnešana tiek veikta absorbēto saules staru enerģijas dēļ.
Šo baktēriju fotosintēzi, kas notiek bez skābekļa izdalīšanās, sauc par fotoreducēšanu. Oglekļa dioksīda fotoreducēšana ir saistīta ar ūdeņraža pārnešanu nevis no ūdens, bet no sērūdeņraža:
6СО 2 +12Н 2 S+hv → С6Н 12 О 6 +12S=6Н 2 О
Ķīmijsintēzes un baktēriju fotosintēzes bioloģiskā nozīme planētu mērogā ir salīdzinoši neliela. Sēra aprites procesā dabā nozīmīgu lomu spēlē tikai ķīmiskās sintētiskās baktērijas. Uzsūcas zaļie augi sērskābes sāļu veidā sērs tiek reducēts un kļūst par olbaltumvielu molekulu sastāvdaļu. Tālāk, kad putrefaktīvās baktērijas iznīcina mirušās augu un dzīvnieku atliekas, sērs izdalās sērūdeņraža veidā, ko sēra baktērijas oksidē par brīvu sēru (vai sērskābi), veidojot augsnē sulfītus, kas ir pieejami augiem. Ķīmiskās un fotoautotrofās baktērijas ir būtiskas slāpekļa un sēra ciklā.
Sporulācija
Sporas veidojas baktēriju šūnas iekšpusē. Sporulācijas procesa laikā baktēriju šūnā notiek vairāki bioķīmiski procesi. Brīvā ūdens daudzums tajā samazinās un fermentatīvā aktivitāte samazinās. Tādējādi tiek nodrošināta sporu noturība pret nelabvēlīgiem vides apstākļiem (augsta temperatūra, augsta sāls koncentrācija, žāvēšana utt.). Sporulācija ir raksturīga tikai nelielai baktēriju grupai.

Strīdi nav nepieciešams posms dzīves cikls baktērijas. Sporulācija sākas tikai ar barības vielu trūkumu vai vielmaiņas produktu uzkrāšanos. Baktērijas sporu veidā ilgstoši var palikt miera stāvoklī. Baktēriju sporas var izturēt ilgstošu vārīšanu un ļoti ilgu sasalšanu. Kad rodas labvēlīgi apstākļi, sporas uzdīgst un kļūst dzīvotspējīgas. Baktēriju sporas ir adaptācija, lai izdzīvotu nelabvēlīgos apstākļos.
Pavairošana
Baktērijas vairojas, sadalot vienu šūnu divās daļās. Sasniedzot noteiktu izmēru, baktērija sadalās divās identiskās baktērijās. Tad katrs no tiem sāk baroties, aug, dalās utt.

Pēc šūnu pagarināšanas pamazām veidojas šķērseniskā starpsiena, un tad atdalās meitas šūnas; Daudzās baktērijās noteiktos apstākļos pēc dalīšanās šūnas paliek savienotas raksturīgās grupās. Šajā gadījumā atkarībā no dalīšanas plaknes virziena un dalījumu skaita rodas dažādas formas. Vairošanās ar pumpuru veidošanu notiek kā izņēmums baktērijām.
Labvēlīgos apstākļos šūnu dalīšanās daudzās baktērijās notiek ik pēc 20-30 minūtēm. Ar tik strauju vairošanos vienas baktērijas pēcnācēji 5 dienās spēj veidot masu, kas var piepildīt visas jūras un okeānus. Vienkāršs aprēķins parāda, ka dienā var izveidoties 72 paaudzes (720 000 000 000 000 000 000 šūnas). Pārrēķinot uz svaru - 4720 tonnas. Tomēr dabā tas nenotiek, jo lielākā daļa baktēriju ātri mirst, saskaroties ar tām saules gaisma, žāvēšanas laikā, barības trūkums, karsēšana līdz 65-100ºС, sugu cīņas rezultātā utt.
Baktērija (1), uzņemot pietiekami daudz pārtikas, palielinās izmērs (2) un sāk gatavoties reprodukcijai (šūnu dalīšanās). Tās DNS (baktērijā DNS molekula ir noslēgta gredzenā) dubultojas (baktērija rada šīs molekulas kopiju). Abas DNS molekulas (3, 4) ir piestiprinātas pie baktērijas sienas un, baktērijai pagarinoties, attālinās (5, 6). Vispirms sadalās nukleotīds, tad citoplazma.

Pēc divu DNS molekulu diverģences uz baktērijas parādās sašaurināšanās, kas pakāpeniski sadala baktērijas ķermeni divās daļās, no kurām katra satur DNS molekulu (7).
Gadās (Bacillus subtilis), ka divas baktērijas salīp kopā un starp tām veidojas tilts (1,2).

Džemperis transportē DNS no vienas baktērijas uz otru (3). Nokļūstot vienā baktērijā, DNS molekulas savijas, dažās vietās salīp kopā (4) un pēc tam apmainās ar sekcijām (5).
Baktēriju loma dabā
Gyre
Baktērijas ir vissvarīgākā saikne vispārējā vielu ciklā dabā. Augi veido sarežģītas organiskas vielas no oglekļa dioksīda, ūdens un minerālsāļiem augsnē. Šīs vielas atgriežas augsnē ar mirušām sēnītēm, augiem un dzīvnieku līķiem. Baktērijas sadala sarežģītas vielas vienkāršās, kuras pēc tam izmanto augi.
Baktērijas iznīcina mirušo augu un dzīvnieku līķu kompleksās organiskās vielas, dzīvo organismu ekskrēcijas un dažādus atkritumus. Barojot ar šīm organiskajām vielām, saprofītiskās sabrukšanas baktērijas pārvērš tās humusā. Tie ir sava veida mūsu planētas sakārtotāji. Tādējādi baktērijas aktīvi piedalās vielu apritē dabā.
Augsnes veidošanās
Tā kā baktērijas ir izplatītas gandrīz visur un sastopamas milzīgā daudzumā, tās lielā mērā nosaka dažādus dabā notiekošos procesus. Rudenī kokiem un krūmiem krīt lapas, iet bojā stiebrzāļu virszemes dzinumi, nobirst vecie zari, ik pa laikam nobirst veco koku stumbri. Tas viss pamazām pārvēršas humusā. 1 cm3. Meža augsnes virskārtā ir simtiem miljonu vairāku sugu saprofītu augsnes baktēriju. Šīs baktērijas pārvērš humusu dažādās minerālvielas, ko no augsnes var uzsūkt augu saknes.
Dažas augsnes baktērijas spēj absorbēt slāpekli no gaisa, izmantojot to dzīvībai svarīgos procesos. Šīs slāpekli fiksējošās baktērijas dzīvo neatkarīgi vai apmetas pākšaugu saknēs. Iekļūstot pākšaugu saknēs, šīs baktērijas izraisa sakņu šūnu augšanu un mezgliņu veidošanos uz tām.
Šīs baktērijas ražo slāpekļa savienojumus, ko izmanto augi. Baktērijas iegūst ogļhidrātus un minerālsāļus no augiem. Tādējādi starp pākšaugu un mezgla baktērijām pastāv cieša saistība, kas ir labvēlīga gan vienam, gan otram organismam. Šo parādību sauc par simbiozi.
Pateicoties simbiozei ar mezgliņu baktērijām, pākšaugi bagātina augsni ar slāpekli, palīdzot palielināt ražu.
Izplatība dabā
Mikroorganismi ir visuresoši. Vienīgie izņēmumi ir aktīvo vulkānu krāteri un nelielas teritorijas sprāgstošas atombumbu epicentros. Ne Antarktīdas zemās temperatūras, ne verdošās geizeru straumes, ne piesātinātie sāls šķīdumi sāls baseinos, ne stiprā kalnu virsotņu insolācija, ne skarbā kodolreaktoru apstarošana netraucē mikrofloras pastāvēšanu un attīstību. Visas dzīvās būtnes pastāvīgi mijiedarbojas ar mikroorganismiem, bieži vien ir ne tikai to krātuves, bet arī izplatītāji. Mikroorganismi ir mūsu planētas vietējie iedzīvotāji, kas aktīvi pēta visneticamākos dabiskos substrātus.
Augsnes mikroflora
Baktēriju skaits augsnē ir ārkārtīgi liels – simtiem miljonu un miljardu īpatņu uz gramu. Augsnē to ir daudz vairāk nekā ūdenī un gaisā. Kopējais baktēriju skaits augsnēs mainās. Baktēriju skaits ir atkarīgs no augsnes veida, to stāvokļa un slāņu dziļuma.
Uz augsnes daļiņu virsmas mikroorganismi atrodas nelielās mikrokolonijās (katrā pa 20-100 šūnām). Tie bieži veidojas organisko vielu recekļu biezumā, uz dzīvām un mirstošām augu saknēm, tievos kapilāros un iekšā kunkuļos.
Augsnes mikroflora ir ļoti daudzveidīga. Šeit ir dažādas baktēriju fizioloģiskās grupas: pūšanas baktērijas, nitrificējošās baktērijas, slāpekli fiksējošās baktērijas, sēra baktērijas uc starp tām ir aerobās un anaerobās, sporu un nesporu formas. Mikroflora ir viens no augsnes veidošanās faktoriem.
Mikroorganismu attīstības zona augsnē ir zona, kas atrodas blakus dzīvo augu saknēm. To sauc par rizosfēru, un tajā esošo mikroorganismu kopumu sauc par rizosfēras mikrofloru.
Rezervuāru mikroflora
Ūdens ir dabiska vide, kurā mikroorganismi attīstās lielā skaitā. Lielākā daļa no tiem nonāk ūdenī no augsnes. Faktors, kas nosaka baktēriju skaitu ūdenī un barības vielu klātbūtni tajā. Tīrākie ūdeņi ir no artēziskajiem akām un avotiem. Atvērtās ūdenskrātuves un upes ir ļoti bagātas ar baktērijām. Lielākais baktēriju skaits ir atrodams ūdens virszemes slāņos, tuvāk krastam. Attālinoties no krasta un palielinoties dziļumam, baktēriju skaits samazinās.
Tīrā ūdenī ir 100-200 baktērijas uz ml, un piesārņotā ūdenī ir 100-300 tūkstoši vai vairāk. Apakšējā dūņās ir daudz baktēriju, īpaši virsmas slānī, kur baktērijas veido plēvi. Šī plēve satur daudz sēra un dzelzs baktēriju, kas oksidē sērūdeņradi līdz sērskābei un tādējādi novērš zivju bojāeju. Dūņos ir vairāk sporu nesošo formu, savukārt ūdenī dominē nesporas formas.
Pēc sugu sastāva ūdens mikroflora ir līdzīga augsnes mikroflorai, taču ir arī specifiskas formas. Iznīcinot dažādus ūdenī nonākušos atkritumus, mikroorganismi pamazām veic tā saukto bioloģisko ūdens attīrīšanu.
Gaisa mikroflora
Gaisa mikroflora ir mazāka nekā augsnes un ūdens mikroflora. Baktērijas paceļas gaisā ar putekļiem, var tur kādu laiku palikt, un pēc tam nosēsties uz zemes virsmas un iet bojā no uztura trūkuma vai ultravioleto staru ietekmē. Mikroorganismu skaits gaisā ir atkarīgs no ģeogrāfiskās zonas, reljefa, gada laika, putekļu piesārņojuma uc katrs putekļu plankums ir mikroorganismu nesējs. Lielākā daļa baktēriju atrodas gaisā virs rūpniecības uzņēmumiem. Laukos gaiss ir tīrāks. Tīrākais gaiss ir virs mežiem, kalniem un sniegotām vietām. Augšējos gaisa slāņos ir mazāk mikrobu. Gaisa mikroflorā ir daudz pigmentētu un sporu saturošu baktēriju, kas ir izturīgākas par citām pret ultravioletajiem stariem.
Cilvēka ķermeņa mikroflora
Cilvēka ķermenis, pat pilnīgi vesels, vienmēr ir mikrofloras nesējs. Cilvēka ķermenim saskaroties ar gaisu un augsni, uz apģērba un ādas nogulsnējas dažādi mikroorganismi, arī patogēnie (stingumkrampju baciļi, gāzes gangrēna u.c.). Atklātās daļas, visticamāk, tiks piesārņotas. cilvēka ķermenis. Uz rokām atrodami E. coli un stafilokoki. Mutes dobumā ir vairāk nekā 100 veidu mikrobu. Mute ar savu temperatūru, mitrumu un barības vielu atliekām ir lieliska vide mikroorganismu attīstībai.
Kuņģī notiek skāba reakcija, tāpēc lielākā daļa tajā esošo mikroorganismu mirst. Sākot no tievās zarnas, reakcija kļūst sārmaina, t.i. labvēlīgs mikrobiem. Mikroflora resnajā zarnā ir ļoti daudzveidīga. Katrs pieaugušais ik dienas ar ekskrementiem izdala aptuveni 18 miljardus baktēriju, t.i. vairāk indivīdu nekā cilvēku uz zemeslodes.
Iekšējos orgānos, kas nav saistīti ar ārējo vidi (smadzenes, sirds, aknas, urīnpūslis utt.), parasti nav mikrobu. Mikrobi šajos orgānos iekļūst tikai slimības laikā.
Baktērijas vielu ciklā
Mikroorganismiem kopumā un jo īpaši baktērijām ir liela nozīme bioloģiski svarīgajos vielu ciklos uz Zemes, veicot ķīmiskas transformācijas, kas ir pilnīgi nepieejamas ne augiem, ne dzīvniekiem. Dažādus elementu cikla posmus veic dažāda veida organismi. Katras atsevišķas organismu grupas pastāvēšana ir atkarīga no elementu ķīmiskās transformācijas, ko veic citas grupas.
Slāpekļa cikls
Slāpekļa savienojumu cikliskajai transformācijai ir galvenā loma nepieciešamo slāpekļa formu apgādē biosfēras organismiem ar dažādām uztura vajadzībām. Vairāk nekā 90% no kopējā slāpekļa fiksācijas notiek dažu baktēriju metaboliskās aktivitātes dēļ.
Oglekļa cikls
Organiskā oglekļa bioloģiskai pārvēršanai oglekļa dioksīdā, ko pavada molekulārā skābekļa samazināšanās, ir nepieciešama dažādu mikroorganismu kopīga vielmaiņas aktivitāte. Daudzas aerobās baktērijas veic pilnīgu organisko vielu oksidāciju. Aerobos apstākļos organiskie savienojumi sākotnēji tiek sadalīti fermentācijas ceļā, un fermentācijas organiskie gala produkti tiek tālāk oksidēti anaerobās elpošanas ceļā, ja ir neorganiskie ūdeņraža akceptori (nitrāti, sulfāti vai CO 2 ).
Sēra cikls
Sērs ir pieejams dzīviem organismiem galvenokārt šķīstošu sulfātu vai reducētu organisko sēra savienojumu veidā.
Dzelzs cikls
Dažās saldūdens tilpnēs ir liela reducēto dzelzs sāļu koncentrācija. Šādās vietās veidojas specifiska baktēriju mikroflora – dzelzs baktērijas, kas oksidē reducēto dzelzi. Tie piedalās purva dzelzsrūdu un ar dzelzs sāļiem bagātu ūdens avotu veidošanā.
Baktērijas ir senākie organismi, kas arhejā parādījās apmēram pirms 3,5 miljardiem gadu. Apmēram 2,5 miljardus gadu viņi dominēja uz Zemes, veidojot biosfēru un piedalījās skābekļa atmosfēras veidošanā.
Baktērijas ir vieni no vienkāršāk strukturētajiem dzīviem organismiem (izņemot vīrusus). Tiek uzskatīts, ka tie ir pirmie organismi, kas parādījās uz Zemes.
Baktēriju šūnas strukturālās sastāvdaļas ir sadalītas 2 veidos:
- pamatstruktūras(šūnu siena, citoplazmas membrāna ar tās atvasinājumiem, citoplazma ar ribosomām un dažādiem ieslēgumiem, nukleoīds);
- pagaidu būves(kapsula, gļotāda, flagellas, bārkstiņas, endosporas, veidojas tikai noteiktos baktēriju dzīves cikla posmos).
Pamatstruktūras.
Šūnapvalki kas atrodas citoplazmas membrānas ārpusē. Citoplazmas membrāna nav daļa no šūnas sienas. Šūnu sienas funkcijas:
Baktēriju aizsardzība pret osmotisko šoku un citiem kaitīgiem faktoriem;
Baktēriju formas noteikšana;
Dalība baktēriju metabolismā.
Šūnu siena ir caurstrāvota ar porām, caur kurām tiek transportēti baktēriju eksotoksīni. Šūnu sienas biezums ir 10–100 nm. Baktēriju šūnu sienas galvenā sastāvdaļa ir peptidoglikāns vai mureīns, kas sastāv no mainīgiem N-acetil-N-glikozamīna un N-acetilmuramīnskābes atlikumiem, kas savienoti ar glikozīdu saitēm.
1884. gadā H. Grams ierosināja metodi baktēriju krāsošanai, izmantojot genciānas violetu, jodu, etilspirtu un fuksīnu. Visas baktērijas atkarībā no to grama traipa ir sadalītas 2 grupās: grampozitīvās un gramnegatīvās baktērijas. Grampozitīvu baktēriju šūnu siena cieši pieguļ citoplazmas membrānai, tās biezums ir 20-100 nm. Tas satur teihoskābes (glicerīna vai ribitola polimērus), kā arī nelielu daudzumu polisaharīdu, proteīnu un lipīdu. Gramnegatīvu baktēriju šūnu siena daudzslāņu, tā biezums ir 14-17 nm. Iekšējais slānis (peptidoglikāns) veido plānu nepārtrauktu tīklu. Ārējais slānis sastāv no fosfolipīdiem, lipoproteīniem un olbaltumvielām. Ārējās membrānas proteīni ir cieši saistīti ar peptidoglikāna slāni.
Noteiktos apstākļos baktērijas zaudē spēju pilnībā vai daļēji sintezēt šūnu sienas komponentus, kā rezultātā veidojas protoplasti, sferoplasti un baktēriju L-formas. Sferoplasti ir baktērijas ar daļēji iznīcinātu šūnu sieniņu. Tos novēro gramnegatīvās baktērijās. Protoplasti- tās ir formas, kurām pilnībā nav šūnu sienas. Tos veido grampozitīvas baktērijas. L-forma baktērijas ir baktēriju mutanti, kas daļēji vai pilnībā zaudējuši spēju sintezēt šūnu sienas peptidoglikānu (baktērijas ar bojātu šūnu sieniņu). Viņi savu vārdu ieguvuši no Listera institūta Anglijā nosaukuma, kur tie tika atvērti 1935. gadā.
Citoplazmas membrāna (CPM) un tās atvasinājumi. Citoplazmas membrāna (plazmolemma) ir baktēriju šūnas daļēji caurlaidīga lipoproteīnu struktūra, kas atdala citoplazmu no šūnas sienas. Tas veido 8-15% no šūnas sausās masas. Tās iznīcināšana izraisa šūnu nāvi. Elektronu mikroskopija atklāja tā trīsslāņu struktūru. Citoplazmas membrāna ir olbaltumvielu (50-75%) un lipīdu (15-20%) komplekss. Lielāko daļu lipīdu veido fosfolipīdi. Turklāt membrānā tika atrasts neliels daudzums ogļhidrātu.
Baktēriju CPM veic šādas funkcijas:
Barjeras funkcija (molekulārais “siets”);
Enerģija;
Selektīva dažādu organisko un neorganisko molekulu un jonu pārnešana, izmantojot īpašus nesējus - translokāzes vai permeāzes;
Replikācija un sekojoša hromosomu dalīšanās.
Šūnu augšanas laikā citoplazmas membrāna veido daudzas invaginācijas (invaginācijas), ko sauc mezosomas.
Citoplazma - Tas ir baktēriju šūnas saturs, ko ierobežo citoplazmas membrāna. Tas sastāv no citozola un strukturālajiem elementiem.
Citozols- homogēna frakcija, ieskaitot šķīstošās RNS sastāvdaļas, fermentus un vielmaiņas produktus.
Strukturālie elementi- tās ir ribosomas, intracitoplazmas membrānas, ieslēgumi un nukleoīds.
Ribosomas- organoīdi, kas veic olbaltumvielu biosintēzi. Tie sastāv no olbaltumvielām un RNS. Tās ir granulas ar diametru 15-20 nm. Viena baktēriju šūna satur no 5000 līdz 50 000 ribosomu. Ribosomas ir olbaltumvielu sintēzes vieta.
Prokariotu citoplazmā ir atrodami dažādi ieslēgumi, kas pārstāv šūnas rezerves vielas. No polisaharīdiem šūnās nogulsnējas glikogēns, ciete un cietei līdzīga viela - granuloze. Polifosfāti ir ietverti granulās, ko sauc volutinisks, vai metahromatisks, graudi.
Nukleoīds ir prokariotu kodols. Tas sastāv no vienas divpavedienu DNS virknes, kas noslēgta gredzenā, ko uzskata par baktēriju hromosomu. Nukleoīdam trūkst kodola apvalka.
Papildus nukleoīdam baktēriju šūnā tika atrasti ekstrahromosomu ģenētiskie elementi - plazmīdas, kas ir mazas apļveida DNS molekulas, kas spēj autonomi replikēties. Plazmīdu loma ir tāda, ka tās kodē papildu īpašības, kas piešķir šūnai priekšrocības noteiktos dzīves apstākļos. Visizplatītākās ir tās plazmīdas, kas nosaka baktēriju rezistences pazīmes pret antibiotikām (R-plazmīdas), enterotoksīnu (Ent-plazmīdas) vai hemolizīnu (Hly-plazmīdas) sintēzi.
UZ pagaidu būves ietver kapsulu, flagellas, pili, baktēriju endosporas.
Kapsula - Tas ir gļotādas slānis virs baktērijas šūnas sienas. Kapsulas viela sastāv no polisaharīdu pavedieniem. Kapsula tiek sintezēta uz citoplazmas membrānas ārējās virsmas un tiek izlaista uz šūnas sienas virsmas noteiktās vietās.
Kapsulas funkcijas:
Kapsulāro antigēnu atrašanās vieta, kas nosaka baktēriju virulenci, antigēnu specifiskumu un imunogenitāti;
Šūnu aizsardzība no mehāniskiem bojājumiem, izžūšanas, toksiskām vielām, fāgu inficēšanās, makroorganisma aizsargfaktoru iedarbības;
Šūnu spēja pievienoties substrātam.
Flagella - Tie ir baktēriju kustības orgāni. Flagellas nav dzīvībai svarīgas struktūras, tāpēc atkarībā no augšanas apstākļiem tās baktērijās var būt vai nebūt. Ziedu skaits un to atrašanās vieta dažādās baktērijās ir atšķirīga. Atkarībā no tā tiek izdalītas šādas kažokādu baktēriju grupas:
- monotrihs– baktērijas ar vienu polāri novietotu karogu;
- amfitrihi– baktērijas ar divām polāri izkārtotām karogiem vai ar karogiem abos galos;
- lofotrihs– baktērijas, kurām vienā šūnas galā ir karogs saišķis;
- peritrichous- baktērijas ar daudzām flagellām, kas atrodas šūnas sānos vai visā tās virsmā.
Flagelu ķīmisko sastāvu attēlo olbaltumvielas flagellīns.
Baktēriju šūnas virsmas struktūras ietver arī villi Un dzēra. Šīs struktūras ir iesaistītas šūnu adsorbcijā uz substrāta (villi, vispārīgie pili) un ģenētiskā materiāla pārneses procesos (seksuālie pili). Tos veido īpašs hidrofobs proteīns pilin.
Dažās baktērijās noteiktos apstākļos veidojas neaktīvās formas, kas nodrošina šūnu ilgstošu izdzīvošanu nelabvēlīgos apstākļos - endosporas. Tie ir izturīgi pret nelabvēlīgiem vides faktoriem.
Sporu atrašanās vieta šūnā:
Centrālā (sibīrijas mēra izraisītājs);
Subtermināls - tuvāk beigām (botulisma izraisītājs);
Terminālis - nūjas galā (stingumkrampju izraisītājs).
Papildus 5 dzīvās dabas valstībām ir vēl divas supervalsts: prokarioti un eikarioti. Tāpēc, ja ņemam vērā baktēriju sistemātisko stāvokli, tas būs šāds:
Kāpēc šie organismi ir klasificēti kā atsevišķs taksons? Lieta tāda, ka baktēriju šūnai ir raksturīgas noteiktas pazīmes, kas atstāj iespaidu uz tās dzīves aktivitāti un mijiedarbību ar citām radībām un cilvēkiem.
Baktēriju atklāšana

Ribosomas ir niecīgas struktūras, kas citoplazmā ir izkaisītas lielā skaitā. To dabu attēlo RNS molekulas. Šīs granulas ir materiāls, pēc kura var noteikt noteikta veida baktēriju attiecības pakāpi un sistemātisko atrašanās vietu. To funkcija ir olbaltumvielu molekulu montāža.
Kapsula
Baktēriju šūnai raksturīga aizsargājošu gļotādu klātbūtne, kuras sastāvu nosaka polisaharīdi vai polipeptīdi. Šādas struktūras sauc par kapsulām. Ir mikro- un makrokapsulas. Šī struktūra veidojas ne visās sugās, bet lielākajā daļā, tas ir, tā nav obligāta.
No kā kapsula aizsargā baktēriju šūnu? No saimnieka antivielu izraisītas fagocitozes, ja baktērija ir patogēna. Vai arī no izžūšanas un kaitīgu vielu iedarbības, ja runājam par citiem veidiem.
Gļotas un ieslēgumi
Arī baktēriju izvēles struktūras. Gļotas jeb glikokalikss ir ķīmiski balstītas uz gļotādas polisaharīdu. To var veidot gan šūnas iekšienē, gan ar ārējiem enzīmiem. Ļoti labi šķīst ūdenī. Mērķis: baktēriju piestiprināšana pie pamatnes – adhēzija.
Ieslēgumi ir dažādas ķīmiskās dabas citoplazmas mikrogranulas. Tie var būt olbaltumvielas, aminoskābes, nukleīnskābes vai polisaharīdi.

Kustības organoīdi
Baktērijas šūnas īpašības izpaužas arī tās kustībā. Šim nolūkam ir klāt flagellas, kuru skaits var būt dažāds (no viena līdz vairākiem simtiem vienā šūnā). Katra flagelluma pamatā ir proteīns flagellīns. Pateicoties elastīgajām kontrakcijām un ritmiskām kustībām no vienas puses uz otru, baktērija var pārvietoties telpā. Karogs ir pievienots citoplazmas membrānai. Atrašanās vieta var arī atšķirties atkarībā no dažādi veidi.
Dzēra
Pat smalkākas par flagellas ir struktūras, kas piedalās:
- piestiprināšana pie pamatnes;
- ūdens-sāls uzturs;
- seksuālā reprodukcija.
Tie sastāv no proteīna pilīna, to skaits vienā šūnā var sasniegt vairākus simtus.
Līdzības ar augu šūnām
Baktēriju un tiem ir viena nenoliedzama līdzība - šūnu sienas klātbūtne. Tomēr, ja augos tas nenoliedzami ir, baktērijās tas nav sastopams visās sugās, tas ir, tā ir izvēles struktūra.
Baktēriju šūnu sienas ķīmiskais sastāvs:
- peptidoglikāna mureīns;
- polisaharīdi;
- lipīdi;
- olbaltumvielas.
Parasti šai struktūrai ir dubultslānis: ārējais un iekšējais. Veic tādas pašas funkcijas kā augi. Saglabā un nosaka nemainīgu ķermeņa formu un nodrošina mehānisku aizsardzību.

Strīds par izglītību
Mēs esam detalizēti aplūkojuši baktēriju šūnas struktūru. Atliek tikai minēt, kā baktērijas var izdzīvot labvēlīgi apstākļi, nezaudējot dzīvotspēju ļoti ilgu laiku.
Viņi to dara, veidojot struktūru, ko sauc par strīdu. Tam nav nekāda sakara ar reprodukciju un tikai aizsargā baktērijas no nelabvēlīgiem apstākļiem. Strīdu forma var būt dažāda. Kad tiek atjaunoti normāli vides apstākļi, sporas sākas un pāraug par aktīvu baktēriju.
Prokariotu organismu šūnai ir sarežģīta, stingri sakārtota struktūra, un tai ir būtiskas ultrastrukturālās organizācijas un ķīmiskais sastāvs.
Baktērijas šūnas strukturālās sastāvdaļas iedala pamata un pagaidu (2. att.). Galvenās struktūras ir: šūnu siena, citoplazmas membrāna ar tās atvasinājumiem, citoplazma ar ribosomām un dažādiem ieslēgumiem, nukleoīds; pagaidu - kapsula, gļotāda, flagellas, bārkstiņas, endosporas, veidojas tikai noteiktos baktēriju dzīves cikla posmos; dažās sugās to nav pilnībā.
Prokariotu šūnā struktūras, kas atrodas ārpus citoplazmas membrānas, sauc par virspusējām (šūnas siena, kapsula, flagellas, bārkstiņas).
Termins "aploksne" pašlaik tiek lietots, lai apzīmētu baktēriju šūnu sienu un kapsulu vai tikai šūnu sienu; citoplazmas membrāna nav apvalka daļa un attiecas uz protoplastu.
Šūnu siena ir svarīgs baktēriju šūnas struktūras elements, kas atrodas starp citoplazmas membrānu un kapsulu; nekapsulārajās baktērijās tā ir ārējā šūnu membrāna. Tas ir obligāts visiem prokariotiem, izņemot mikoplazmas un L formas baktērijas. Veic vairākas funkcijas: aizsargā baktērijas no osmotiskā šoka un citiem kaitīgiem faktoriem, nosaka to formu, piedalās vielmaiņā; daudzos patogēno baktēriju veidos tas ir toksisks, satur virsmas antigēnus, kā arī nes uz virsmas specifiskus fāgu receptorus. Baktēriju šūnu sieniņā ir poras, kas ir iesaistītas eksotoksīnu un citu baktēriju eksoproteīnu transportēšanā. Šūnas sienas biezums ir 10–100 nm, un tas veido 5–50% no šūnas sausnas.
Baktēriju šūnas sienas galvenā sastāvdaļa ir peptidoglikāns jeb mureīns (latīņu murus — siena), atbalsta polimērs, kam ir tīklveida struktūra un kas veido stingru (cietu) baktēriju šūnas ārējo karkasu. Peptidoglikānam ir galvenā ķēde (mugurkauls), kas sastāv no mainīgiem N-acetil-M-glikozamīna un N-acetilmuramīnskābes atlikumiem, kas savienoti ar 1,4-glikozīdu saitēm, identiskām tetrapeptīdu sānu ķēdēm, kas pievienotas N-acetilmuramīnskābes molekulām, un īsiem krusteniskiem peptīdiem. ķēdes tilti, kas savieno polisaharīdu ķēdes. Divu veidu saites (glikozīdu un peptīdu), kas savieno peptidoglikāna apakšvienības, piešķir šim heteropolimēram molekulārā tīkla struktūru. Peptidoglikāna slāņa kodols ir vienāds visām baktēriju sugām; Tetrapeptīdu olbaltumvielu ķēdes un peptīdu (šķērsvirziena) ķēdes dažādās sugās atšķiras.
Pamatojoties uz to tinctoric īpašībām, visas baktērijas ir sadalītas divās grupās: grampozitīvās un gramnegatīvās. 1884. gadā H. Grams ierosināja krāsošanas metodi, ko izmantoja baktēriju diferencēšanai. Metodes būtība ir tāda, ka grampozitīvās baktērijas stingri nostiprina genciānas vijolītes un joda kompleksu, nav pakļautas balināšanai ar etanolu un tāpēc neuztver papildu krāsvielu fuksīnu, paliekot purpursarkanā krāsā. Gramnegatīvajās baktērijās šo kompleksu viegli izskalo no šūnas etanols, un, papildus uzklājot fuksīnu, tās kļūst sarkanas. Dažām baktērijām pozitīvs Grama traips tiek novērots tikai aktīvās augšanas stadijā. Prokariotu spēju krāsot ar gramu vai atkrāsot ar etanolu nosaka to šūnu sienas īpašais ķīmiskais sastāvs un ultrastruktūra. Peptidoglikāns grampozitīvās baktērijās ir galvenā šūnu sienas sastāvdaļa un veido no 50 līdz 90%, gramnegatīvās baktērijās tas ir 1-10%. Gramnegatīvo baktēriju peptidoglikāna strukturālās mikrofibrillas ir mazāk kompakti šķērssaites, tāpēc to peptidoglikāna slānī poras ir daudz plašākas nekā grampozitīvo baktēriju molekulārajā karkasā. Ar šādu peptidoglikāna strukturālo organizāciju genciānas violets un joda komplekss gramnegatīvajās baktērijās tiks izskalots ātrāk.
Grampozitīvo baktēriju šūnu siena atrodas cieši blakus citoplazmas membrānai, masīva, un tās biezums ir 20-100 nm robežās. To raksturo teikoskābju klātbūtne, tās ir saistītas ar peptidoglikānu un ir trīsvērtīgā spirta - glicerīna vai pentaatomiskā spirta - ribitola polimēri, kuru atlikumus savieno fosfodiestera saites. Teihoskābes saista magnija jonus un piedalās to transportēšanā šūnā. Polisaharīdi, proteīni un lipīdi nelielos daudzumos ir atrodami arī grampozitīvu prokariotu šūnu sieniņās.
Rīsi. 2. Prokariotu šūnas uzbūves shēma:
1 - kapsula; 2 - šūnu siena; 3 - citoplazmas membrāna; 4 - nukleoīds; 5 - citoplazma; 6 - hromatofori; 7 - tilakoīdi; 8 - mezosoma; 9 - ribosomas; 10 - flagellas; 11-bazālais ķermenis; 12 - dzēra; 13 - sēra iekļaušana; 14 — tauku pilieni; 15 — polifosfāta granulas; 16 - plazmīda
Gramnegatīvo baktēriju šūnu siena ir daudzslāņaina, tās biezums ir 14-17 nm. Iekšējais slānis ir peptidoglikāns, kas veido plānu (2 nm) nepārtrauktu tīklu, kas ieskauj šūnu. Peptidoglikāns satur tikai mezodiaminopimelskābi un nesatur lizīnu. Šūnu sienas ārējais slānis - ārējā membrāna - sastāv no fosfolipīdiem, lipopolisaharīdiem, lipoproteīniem un olbaltumvielām. Ārējā membrāna satur matricas proteīnus, kas ir cieši saistīti ar peptidoglikāna slāni. Viena no to funkcijām ir hidrofilu poru veidošanās membrānā, caur kurām notiek molekulu difūzija ar masu līdz 600, dažreiz 900. Matricas proteīni turklāt darbojas arī kā receptori dažiem fāgiem. Lipopolisaharīds (LPS) gramnegatīvo baktēriju šūnu sieniņās sastāv no lipīda A un polisaharīda. LPS, kas ir toksisks dzīvniekiem, sauc par endotoksīnu. Teihoskābes nav atrastas gramnegatīvās baktērijās.
Gramnegatīvo baktēriju šūnu sienas strukturālās sastāvdaļas ir norobežotas no citoplazmas membrānas un atdalītas ar telpu, ko sauc par periplazmu vai periplazmas telpu.
Protoplasti un sferoplasti. Protoplasti ir prokariotu formas, kurām pilnībā nav šūnu sienas, parasti veidojas grampozitīvās baktērijās. Sferoplasti ir baktērijas ar daļēji iznīcinātu šūnu sieniņu. Tie saglabā ārējās membrānas elementus. Tos novēro gramnegatīvās baktērijās un daudz retāk grampozitīvās baktērijās. Tie veidojas peptidoglikāna slāņa iznīcināšanas rezultātā ar lītiskiem enzīmiem, piemēram, lizocīmu, vai bloķējot peptidoglikāna biosintēzi ar antibiotiku penicilīnu utt. vidē ar atbilstošu osmotisko spiedienu.
Protoplastiem un sferoplastiem ir sfēriska vai puslodes forma, un tie ir 3-10 reizes lielāki nekā sākotnējās šūnas. Normālos apstākļos notiek osmotiskā līze un viņi mirst. Paaugstināta osmotiskā spiediena apstākļos tie kādu laiku spēj izdzīvot, augt un pat dalīties. Kad tiek noņemts faktors, kas iznīcina peptidoglikānu, protoplasti, kā likums, mirst, bet var pārvērsties L formās; sferoplasti viegli atgriežas pie sākotnējām baktērijām, dažreiz pārvēršas L formās vai mirst.
L-baktēriju formas. Tās ir baktēriju fenotipiskas modifikācijas jeb mutanti, kas daļēji vai pilnībā zaudējuši spēju sintezēt šūnu sienas peptidoglikānu. Tādējādi L-formas ir baktērijas, kurām ir defekti šūnu sieniņā. Savu nosaukumu viņi ieguvuši tāpēc, ka 1935. gadā tika izolēti un aprakstīti Listera institūtā Anglijā. Tie veidojas L-transformējošo aģentu – antibiotiku (penicilīns, polimiksīns, bacitracīns, venkomicīns, streptomicīns), aminoskābju ietekmē. (glicīns, metionīns, leicīns utt.), enzīms lizocīms, ultravioletie un rentgena stari. Atšķirībā no protoplastiem un sferoplastiem, L-formām ir salīdzinoši augsta dzīvotspēja un izteikta spēja vairoties. Morfoloģisko un kultūras īpašību ziņā tās krasi atšķiras no sākotnējām baktērijām, kas ir saistīts ar šūnu sienas zudumu un vielmaiņas aktivitātes izmaiņām.
Baktēriju L-formas ir polimorfas. Ir elementārie ķermeņi ar izmēru 0,2-1 mikroni (minimāli reproduktīvie elementi), sfēras - 1-5, lieli ķermeņi - 5-50, pavedieni - līdz 4 mikroniem vai vairāk. L formas šūnām ir labi attīstīta intracitoplazmas membrānu sistēma un mielīnam līdzīgas struktūras. Šūnu sieniņas defekta dēļ tie ir osmotiski nestabili, un tos var kultivēt tikai īpašās barotnēs ar augstu osmotisko spiedienu; tie iziet cauri baktēriju filtriem.
Pastāv stabilas un nestabilas baktēriju L formas. Pirmajiem pilnībā nav stingras šūnu sienas, kas padara tos līdzīgus protoplastiem; tie ārkārtīgi reti atgriežas sākotnējās baktēriju formās. Pēdējiem var būt šūnu sienas elementi, kuros tie ir līdzīgi sferoplastiem; ja nav faktora, kas izraisīja to veidošanos, tie tiek atgriezti sākotnējās šūnās.
L-formu veidošanās procesu sauc par L-transformāciju vai L-indukciju. Gandrīz visu veidu baktērijām, ieskaitot patogēnās (brucelozes, tuberkulozes, listērijas uc izraisītājus), ir iespēja pakļaut L-transformāciju.
L-formām tiek piešķirta liela nozīme hronisku recidivējošu infekciju attīstībā, patogēnu pārnēsāšanā un ilgstošā noturībā organismā. Ir pierādīta L formas baktēriju elementāro ķermeņu transplacentālā invazivitāte.
Infekcijas procesam, ko izraisa baktēriju L-formas, ir raksturīga netipiskums, gaitas ilgums, slimības smagums, un to ir grūti ārstēt ar ķīmijterapiju.
Kapsula ir gļotādas slānis, kas atrodas virs baktērijas šūnas sienas. Kapsulas viela ir skaidri norobežota no vidi. Atkarībā no slāņa biezuma un savienojuma stipruma ar baktēriju šūnu makrokapsula, kuras biezums ir lielāks par 0,2 mikroniem, skaidri redzama gaismas mikroskopā, un mikrokapsula, kuras biezums ir mazāks par 0,2 mikroniem, nosakāms tikai ar elektronu mikroskopu vai atklāti ar ķīmiskām un imunoloģiskām metodēm, izšķir. Makrokapsulu (īsto kapsulu) veido B. anlhracis, C1. perfringens, mikrokapsula - Escherichia coJi. Kapsula nav būtiska baktēriju šūnas struktūra: tās zudums neizraisa baktērijas nāvi. Ir zināmi baktēriju mutanti bez kapsulām, piemēram, Sibīrijas mēra vakcīnas celms STI-1.
Kapsulu viela sastāv no ļoti hidrofilām micellām, un to ķīmiskais sastāvs ir ļoti daudzveidīgs. Lielākajai daļai prokariotu kapsulu galvenās sastāvdaļas ir homo- vai hetsropolisaharīdi (entsrobaktērijas utt.). Dažos baciļu veidos kapsulas ir izgatavotas no polipeptīda. Tādējādi B. anthracis kapsulas sastāvs ietver D-glutamīnskābes polipeptīdu (pa labi rotējošais izomērs). Zīdītāju Mycobacterium tuberculosis mikrokapsulas sastāvā ietilpst glikopeptīdi, ko attēlo trehalozes un mikolskābes (nabassaites faktora) esteris.
Kapsulas sintēze ir sarežģīts process, un tam ir savas īpašības dažādos prokariotos; Tiek uzskatīts, ka kapsulas biopolimēri tiek sintezēti uz citoplazmas membrānas ārējās virsmas un izdalās uz šūnas sienas virsmas noteiktos specifiskos apgabalos.
Ir baktērijas, kas sintezē gļotas, kas nogulsnējas uz šūnas sienas virsmas bezstrukturāla polisaharīda rakstura slāņa veidā. Gļotāda, kas ieskauj šūnu, bieži ir biezāka par šūnas diametru. Saprofītiskajā baktērijā Leuconostoca tiek novērota vienas kapsulas veidošanās daudziem indivīdiem. Šādas baktēriju uzkrāšanās, kas atrodas kopējā kapsulā, sauc par zooglea.
Kapsula ir daudzfunkcionāla organelle, kurai ir svarīga bioloģiskā loma. Tā ir kapsulāro antigēnu lokalizācijas vieta, kas nosaka baktēriju virulenci, antigēnu specifiskumu un imunogenitāti. Kapsulas zudums patogēnās baktērijās krasi samazina to virulenci, piemēram, Sibīrijas mēra baciļu nekapsulāros celmos. Kapsulas nodrošina baktēriju izdzīvošanu, pasargājot tās no mehāniskiem bojājumiem, izžūšanas, inficēšanās ar fāgiem, toksiskām vielām, bet patogēnās formās - no makroorganisma aizsargspēku darbības: iekapsulētās šūnas ir vāji fagocitētas. Dažos baktēriju veidos, tostarp patogēnās, tas veicina šūnu piesaisti substrātam.
Veterinārajā mikrobioloģijā kapsulas noteikšana tiek izmantota kā patogēna diferenciāla morfoloģiskā pazīme, pārbaudot uz Sibīrijas mēri.
Kapsulu krāsošanai tiek izmantotas īpašas metodes - Romanovskis - Giemsa, Gins - Burri, Olts, Mikhins utt.
Mikrokapsulas un gļotādas slāni nosaka seroloģiskās reakcijas (RA), kapsulas antigēnās sastāvdaļas tiek identificētas, izmantojot imūnfluorescences metodi (RIF) un RDD.
Flagellas ir baktēriju kustības organellas, kuras attēlo plānas, garas, pavedienam līdzīgas proteīna struktūras. To garums vairākas reizes pārsniedz baktēriju šūnu un ir 10-20 mikroni, un dažās spirilās tas sasniedz 80-90 mikronus. Flageluma pavediens (fibrila) ir pilnīgs spirālveida cilindrs ar diametru 12-20 nm. Vibrios un Proteus kvēldiegu ieskauj 35 nm biezs apvalks.
Žogs sastāv no trim daļām: spirālveida pavediena, āķa un pamata korpusa. Āķis ir izliekts proteīna cilindrs, kas darbojas kā elastīga saite starp bazālo korpusu un stingro kauliņa pavedienu. Bāzes korpuss ir sarežģīta struktūra, kas sastāv no centrālā stieņa (ass) un gredzeniem.

Rīsi. 3. Karogs:
a - monotrihs; b - amfitrihs; c - lofotrihs; d - peritrichous
Flagellas nav būtiskas baktēriju šūnas struktūras: baktērijās ir fāzes variācijas, kad tās atrodas vienā šūnu attīstības fāzē un nav citā. Tādējādi vecajās kultūrās stingumkrampju izraisītājā dominē šūnas bez flagellas.
Ziedu skaits (no I līdz 50 un vairāk) un to lokalizācijas vietas dažādu sugu baktērijās nav vienādas, bet ir stabilas vienai sugai. Atkarībā no tā tiek izdalītas šādas flagellatu baktēriju grupas: moiotrichs - baktērijas ar vienu polāri novietotu flagellum; amphitrichous - baktērijas ar divām polāri izkārtotām karogiem vai kam abos galos ir kašķa saišķis; lophotrichs - baktērijas ar karogs saišķi vienā šūnas galā; peritrichs ir baktērijas ar daudzām flagellām, kas atrodas šūnas sānos vai visā tās virsmā (3. att.). Baktērijas, kurām nav flagellas, sauc par atrihiju.
Kā kustības orgāni, flagellas ir raksturīgas peldošām stieņveida un vītņotām baktēriju formām un tikai atsevišķos gadījumos ir sastopamas kokcīšos. Tie nodrošina efektīvu kustību šķidrā vidē un lēnāku kustību uz cietu pamatņu virsmas. Monotrihu un lofotrihu kustības ātrums sasniedz 50 μm/s, amfitrihs un peritrihs pārvietojas lēnāk un parasti 1 s laikā veic attālumu, kas vienāds ar savas šūnas lielumu.
Baktērijas pārvietojas nejauši, bet tās spēj virzītas kustības formas - taksometri, ko nosaka ārējie stimuli. Reaģējot uz dažādiem vides faktoriem, baktērijas īsā laikā tiek lokalizētas optimālā biotopa zonā. Taksometri var būt pozitīvi un negatīvi. Ir ierasts atšķirt: ķemotaksi, aerotaksi, fototaksi, magnotaksi. Ķīmijtaksis izraisa ķīmisko vielu koncentrācijas atšķirības vidē, aerotakss ar skābekli, fototaksis pēc gaismas intensitātes, magnetotaksi nosaka mikroorganismu spēja orientēties magnētiskajā laukā.
Baktēriju kustīgo karogveidīgo formu identificēšana ir svarīga to identificēšanai infekcijas slimību laboratoriskajā diagnostikā.
Pili (fimbriae, villi) ir taisni, plāni, dobi proteīna cilindri, kuru biezums ir 3-25 nm un garums līdz 12 µm, kas stiepjas no baktēriju šūnas virsmas. Tos veido specifisks proteīns - pilīns, cēlušies no citoplazmas membrānas, atrodami kustīgās un nekustīgās baktēriju formās un ir redzamas tikai elektronu mikroskopā (4. att.). Uz šūnas virsmas var būt no 1-2, 50-400 vai vairāk pili līdz vairākiem tūkstošiem.

Rīsi. 4. Dzēra
Ir divas pilu klases: seksuālie pili (sexpili) un vispārīgie pili, ko biežāk sauc par fimbrijām. Vienai un tai pašai baktērijai var būt dažāda rakstura pili. Dzimuma pilieni parādās uz baktēriju virsmas konjugācijas procesa laikā un veic organellu funkciju, caur kurām ģenētiskais materiāls (DNS) tiek pārnests no donora uz recipientu.
Vispārējā tipa pilīši atrodas peritrihiāli (Escherichia coli) vai pie poliem (pseudomonas); viena baktērija var saturēt simtiem to. Tie piedalās baktēriju adhēzijā aglomerātos, mikrobu piesaistē dažādiem substrātiem, tostarp šūnām (adhezīvā funkcija), metabolītu transportēšanā, kā arī veicina plēvju veidošanos uz šķidrās vides virsmas; izraisīt sarkano asins šūnu aglutināciju.
Citoplazmas membrāna un tās atvasinājumi. Citoplazmas membrāna (plazmolemma) ir baktēriju šūnu daļēji caurlaidīga lipoproteīnu struktūra, kas atdala citoplazmu no šūnas sienas. Tā ir obligāta daudzfunkcionāla šūnas sastāvdaļa un veido 8-15% no tās sausās masas. Citoplazmas membrānas iznīcināšana noved pie baktēriju šūnas nāves. Ultraplānās sekcijas elektronu mikroskopā atklāj tā trīsslāņu struktūru - divus ierobežojošus osmiofīlos slāņus, katrs 2-3 nm biezs, un vienu osmiofobisku centrālo slāni 4-5 nm biezumā.
Ķīmiski citoplazmas membrāna ir proteīna-lipīdu komplekss, kas sastāv no 50-75% olbaltumvielu un 15-50% lipīdu. Lielāko daļu membrānas lipīdu (70-90%) veido fosfolipīdi. Tas ir veidots no diviem monomolekulāriem proteīnu slāņiem, starp kuriem ir lipīdu slānis, kas sastāv no divām regulāri orientētu lipīdu molekulu rindām.
Citoplazmatiskā membrāna kalpo kā osmotiska barjera šūnai, kontrolē barības vielu ieplūdi šūnā un vielmaiņas produktu izdalīšanos uz āru, satur substrātam raksturīgus permeāzes enzīmus, kas veic aktīvu selektīvu organisko un neorganisko molekulu pārnesi.
Citoplazmas membrānas enzīmi katalizē pēdējos posmus membrānas lipīdu, šūnu sienas komponentu, kapsulas un eksoenzīmu sintēzē; Oksidatīvās fosforilēšanas enzīmi un elektronu transporta enzīmi, kas ir atbildīgi par enerģijas sintēzi, ir lokalizēti uz membrānas.
Šūnu augšanas laikā citoplazmas membrāna veido daudzus invaginātus, kas veido intracitoplazmas membrānas struktūras. Vietējās membrānas invaginācijas sauc par mezosomām. Šīs struktūras ir labi izteiktas grampozitīvās baktērijās, sliktāk izteiktas gramnegatīvās baktērijās un slikti izteiktas riketsijā un mikoplazmās.
Ir izveidota saikne starp mezosomām un baktēriju hromosomu; šādas struktūras sauc par nukleoidosomām. Mezosomas, kas integrētas ar nukleoīdu, piedalās kariokinēzē un citokinēzē mikrobu šūnas, nodrošinot genoma sadalījumu pēc DNS replikācijas beigām un sekojošas meitas hromosomu diverģences. Mezosomas, tāpat kā citoplazmas membrāna, ir baktēriju elpošanas aktivitātes centri, tāpēc tos dažreiz sauc par mitohondriju analogiem. Tomēr mezosomu nozīme vēl nav pilnībā noskaidrota. Tie palielina membrānu darba virsmu, iespējams, pildot tikai strukturālu funkciju, sadalot baktēriju šūnu salīdzinoši atsevišķos nodalījumos, kas rada labvēlīgākus apstākļus fermentatīvo procesu norisei. Patogēnajās baktērijās tie nodrošina eksotoksīnu olbaltumvielu molekulu transportēšanu.
Citoplazma ir baktēriju šūnas saturs, ko ierobežo citoplazmas membrāna. Tas sastāv no citozola - viendabīgas frakcijas, ieskaitot šķīstošās RNS sastāvdaļas, substrātu vielas, fermentus, vielmaiņas produktus un strukturālos elementus - ribosomas, intracitoplazmas membrānas, ieslēgumus un nukleoīdu.
Ribosomas ir organellas, kas veic olbaltumvielu biosintēzi. Tie sastāv no olbaltumvielām un RNS, kas savienoti kompleksā ar ūdeņraža un hidrofobām saitēm. Baktēriju ribosomas ir granulas ar diametru 15-20 nm, sedimentācijas konstante ir 70S, un tās veidojas no divām ribonukleoproteīna apakšvienībām: 30S un 50S. Viena baktēriju šūna var saturēt no 5000 līdz 50 000 ribosomu; caur mRNS tās tiek apvienotas polisomu agregātos, kas sastāv no 50–55 ribosomām ar augstu proteīnu sintēzes aktivitāti.
Baktēriju citoplazmā tiek konstatēti dažāda veida ieslēgumi. Tās var būt cietas, šķidras vai gāzveida, ar proteīna membrānu vai bez tās, un tās neatrodas pastāvīgi. Ievērojama daļa no tiem ir rezerves barības vielas un šūnu vielmaiņas produkti. Rezerves barības vielas ir: polisaharīdi, lipīdi, polifosfāti, sēra nogulsnes uc No polisaharīda rakstura ieslēgumiem visbiežāk sastopams glikogēns un cietei līdzīgā viela granulosa, kas kalpo kā oglekļa un enerģijas avots. Lipīdi šūnās uzkrājas granulu un tauku pilienu veidā, tostarp ar membrānu ieskautas poli-/3-hidroksisviestskābes granulas, kas strauji lauž gaismu un ir skaidri redzamas gaismas mikroskopā. Tiek konstatēti arī Sibīrijas mēra baciļi un aerobās sporas veidojošās saprofītiskās baktērijas. Mikobaktērijas uzkrāj vaskus kā rezerves vielas. Dažu masalu nonbaktēriju, spirilla un citu šūnas satur volutīna granulas, ko veido polifosfāti. Viņiem ir raksturīga metahromāzija: toluidīna zilā un metilēnzilā krāsa tos iekrāso violeti sarkanā krāsā. Volutin granulas spēlē fosfātu noliktavas lomu.
Ieslēgumi, ko ieskauj membrāna, ietver arī gāzu vakuolus vai aerosomas; tie samazina šūnu īpatnējo svaru un ir atrodami ūdens prokariotos.
Nukleoīds ir prokariotu kodols. Tas sastāv no vienas divpavedienu DNS virknes, kas noslēgta 1,1–1,6 nm garā gredzenā, ko uzskata par vienu baktēriju hromosomu jeb genoforu.
Prokariotu nukleoīds no pārējās šūnas nav norobežots ar membrānu - tam trūkst kodola apvalka.
Nukleoīdu struktūrās ietilpst RNS polimerāze, pamata proteīni un trūkst histonu; hromosoma ir noenkurota uz citoplazmas membrānas, bet grampozitīvās baktērijās - uz mezosomām. Baktēriju hromosoma replikējas polikonservatīvā veidā: sākotnējā DNS dubultā spirāle atritinās un katras polinukleotīdu ķēdes veidnē tiek samontēta jauna komplementāra ķēde. Nukleoīdam nav mitotiskā aparāta, un meitas kodolu atdalīšanu nodrošina citoplazmas membrānas augšana.
Baktēriju kodols ir diferencēta struktūra. Atkarībā no šūnu attīstības stadijas nukleoīds var būt diskrēts (pārtraukts) un sastāvēt no atsevišķiem fragmentiem. Tas ir saistīts ar faktu, ka baktēriju šūnas sadalīšanās laikā notiek pēc DNS molekulas replikācijas cikla pabeigšanas un meitas hromosomu veidošanās.
Nukleoīds satur lielāko daļu baktēriju šūnas ģenētiskās informācijas.
Papildus nukleoīdam daudzu baktēriju šūnās ir atrodami ekstrahromosomālie ģenētiskie elementi - plazmīdas, kas ir mazas cirkulāras DNS molekulas, kas spēj autonomi replikēties.