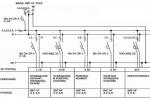
細菌の細胞構造
細菌の構造は、細胞全体とその極薄切片の電子顕微鏡検査を使用してよく研究されています。 細菌細胞は、細胞壁、細胞質膜、封入体を含む細胞質、および核様体と呼ばれる核で構成されています。 さらに、カプセル、マイクロカプセル、粘液、鞭毛、線毛などの構造もあります (図 1)。 一部の細菌は、不利な条件下で胞子を形成することができます。
細胞壁 -細菌に特定の形状を与え、その下にある細胞質膜とともに細菌内の高い浸透圧を「抑制」する、強力で弾性のある構造。 細菌細胞。 細胞分裂および代謝産物の輸送のプロセスに関与しています。 最も厚い細胞壁はグラム陽性菌に見られます (図 1)。 したがって、グラム陰性菌の細胞壁の厚さが約 15 ~ 20 nm である場合、グラム陽性菌では 50 nm 以上に達する可能性があります。 グラム陽性菌の細胞壁には、少量の多糖類、脂質、タンパク質が含まれています。
これらの細菌の細胞壁の主成分は多層構造です。 ペプチドグリカン(ムレイン、ムコペプチド)、細胞壁の質量の 40 ~ 90% を構成します。
ボルチンメソソーマ核様体
米。 1. 細菌細胞の構造。
テイコ酸(ギリシャ語に由来。 テイチョス -壁)、その分子はリン酸架橋によって接続された8〜50個のグリセロール残基とリビトール残基の鎖です。 細菌の形状と強度は、多層でペプチドで架橋されたペプチドグリカンの硬い繊維構造によって与えられます。 ペプチドグリカンは、反復残基からなる並列グリカン分子で表されます。 N-アセチルグルコサミンと N-P 型グリコシド結合で結合したアセチルムラミン酸 (1 -> 4).
リゾチームはアセチルムラミダーゼであり、これらの結合を切断します。 グリカン分子はペプチド架橋によって結合されています。 したがって、このポリマーの名前はペプチドグリカンです。 グラム陰性菌のペプチドグリカンのペプチド結合の基礎は、交互に並んだテトラペプチドです。 L-そして D-アミノ酸。
U 大腸菌ペプチド鎖は互いにつながっています D-一方の鎖はアラニン、もう一方の鎖はメソジアミノピメリン酸です。
グラム陰性菌のペプチドグリカンのペプチド部分の組成と構造は安定していますが、グラム陽性菌のペプチドグリカンはアミノ酸の組成や配列が異なる場合があります。 ここでのテトラペプチドは、5 つのグリシン残基のポリペプチド鎖によって互いに接続されています。 グラム陽性菌には、メソジアミノピメリン酸の代わりにリジンが含まれることがよくあります。 リン脂質

米。 2. グラム陽性菌 (gram+) およびグラム陰性菌 (gram") の表面構造の構造。
グリカン要素 (アセチルグルコサミンとアセチルムラミン酸) およびテトラペプチドアミノ酸 (メソジアミノピメリン酸と L-グルタミン酸、D-アラニン) は、動物や人間には存在しないため、細菌の独特の特徴です。
グラム陽性細菌がグラム染色(細菌の青紫色)を使用して染色した場合にヨウ素と組み合わせてリンドウ紫を保持する能力は、色素と相互作用する多層ペプチドグリカンの特性と関連しています。 さらに、細菌塗抹標本をその後アルコールで処理すると、ペプチドグリカンの細孔が狭くなり、細胞壁に色素が保持されます。 アルコールにさらされると、グラム陰性菌は色素を失って変色し、マゼンタで処理すると赤くなります。 これは、ペプチドグリカンの量が少ないためです (細胞壁質量の 5 ~ 10%)。
グラム陰性菌の細胞壁には、 外膜、リポタンパク質を介してペプチドグリカンの下層に接続されています(図2)。 外膜は内膜と同様に波状の三層構造になっており、細胞質と呼ばれます。 これらの膜の主成分は脂質の二分子(二重)層です。
外膜は、次のように表される非対称モザイク構造です。 リポ多糖類、リン脂質、タンパク質 。 その外側には、 リポ多糖類(LPS)、脂質という3つの成分から構成されています。 あ、コア部分、またはコア(緯度) 芯 -コア)、およびオリゴ糖配列の繰り返しによって形成される 0 特異的多糖鎖。
リポ多糖は脂質によって外膜に「固定」されています あ、 LPS の毒性の原因であることが確認されています。 エンドトキシンあり。 抗生物質による細菌の破壊により大量の内毒素が放出され、患者に内毒素ショックを引き起こす可能性があります。
脂質から あ LPS のコアまたはコア部分が外れます。 LPS コアの最も不変な部分はケトデオキシオクトン酸 (3-デオキシ-g)-マンノ-2-オクツロソン酸) です。 0 -LPS分子のコア部分から伸びる特定の鎖が決定する 血清群、血清型(免疫血清を使用して検出される細菌の一種) 特定の細菌株。 したがって、LPS の概念は、細菌を区別するために使用できる 0 抗原の概念と関連付けられています。 遺伝子の変化は成分の生合成の変化を引き起こす可能性がある LPS細菌とその結果生じるもの L-フォーム
マトリックスタンパク質外膜は、タンパク質分子が ポリナミ、境界には親水性の細孔があり、水と相対質量が 700 までの小さな分子が通過し、外膜と細胞質膜の間には酵素を含むペリプラズム空間 (ペリプラズム) があります。 リゾチーム、ペニシリン、身体の保護因子、その他の化合物の影響で細菌の細胞壁の合成が妨害されると、形状が変化した(多くの場合球形の)細胞が形成されます。 プロトプラスト -細胞壁を完全に欠いている細菌。 スフェロプラスト -細胞壁が部分的に保存された細菌。 細胞壁阻害剤の除去後、そのような変化した細菌は元に戻る可能性があります。 完全な細胞壁を獲得し、元の形状を復元します。
抗生物質などの影響でペプチドグリカンを合成する能力を失い、増殖する能力を持ったスフェロ型またはプロトプラスト型の細菌を細菌と呼びます。 L字型(リスター研究所の名前から)。 L-フォームは突然変異の結果として発生することもあります。 これらは浸透圧に敏感な球形のフラスコ状の細胞で、さまざまなサイズがあり、細菌フィルターを通過する細胞も含まれます。 いくつかの L- 細菌の変化を引き起こした要因が除去されると、形態 (不安定) が逆転し、元の細菌細胞に「戻る」可能性があります。 L- 感染症の多くの病原体によって型が形成される可能性があります。
細胞質膜超薄切片の電子顕微鏡観察では、これは細菌の細胞質の外側部分を取り囲む 3 層の膜です。 構造的には、動物細胞の原形質膜に似ており、主に表面が埋め込まれたリン脂質と膜の構造を貫通しているように見える内在性タンパク質の二重層の脂質で構成されています。 それらの中には、物質の輸送に関与する透過物もあります。 細胞質膜は可動性の構成要素を含む動的構造であるため、可動性の流体構造と考えられています。 浸透圧の調節、物質の輸送、細胞のエネルギー代謝(電子伝達系の酵素、アデノシントリホスファターゼなどによる)に関与しています。 (細胞壁の成長と比較して)過度の成長により、細胞質膜は陥入を形成します。これは、複雑にねじれた膜構造の形で陥入を形成し、 メソソーム。それほど複雑ではないねじれた構造は次のように呼ばれます。 細胞質内膜。メソソームと細胞質内膜の役割は完全には理解されていません。 これらは、電子顕微鏡用に標本を準備(固定)した後に発生するアーチファクトであるとさえ示唆されています。 それにもかかわらず、細胞質膜の誘導体は細胞分裂に参加し、細胞壁の合成にエネルギーを提供し、物質の分泌、胞子形成、すなわち細胞の形成に関与すると考えられています。 エネルギー消費量の多いプロセスで。
細胞質細菌細胞の大部分を占め、可溶性タンパク質、リボ核酸、封入体、および多数の小さな顆粒で構成されています。 リボソームタンパク質の合成(翻訳)を担当します。 細菌のリボソームは約 20 nm のサイズと沈降係数を持っています。 70年代、 3 真核細胞に特徴的な 80^-リボソームとの違い。 したがって、一部の抗生物質は、細菌のリボソームに結合することにより、真核細胞のタンパク質合成に影響を与えることなく、細菌のタンパク質合成を抑制します。 細菌のリボソームは 2 つのサブユニットに解離することがあります。 50年代そして 30代 . 細胞質には、グリコーゲン顆粒、多糖類、ポリ-p-酪酸、ポリリン酸塩(ボルチン)などのさまざまな封入体が含まれています。 それらは環境中に過剰な栄養素がある場合に蓄積し、栄養とエネルギーの必要性に対する予備物質として機能します。 ボルチンは塩基性色素に親和性があり、異色症を示し、特別な染色法を使用すると簡単に検出されます。 ジフテリア桿菌では、ボルチン粒子の特徴的な配置が、強く染色された細胞極の形で明らかになります。
核様体 - 細菌の核に相当します。 それは二本鎖DNAの形で細菌の中央領域に位置し、リングで閉じられ、ボールのように密集しています。 真核生物とは異なり、細菌の核には核膜、核小体、塩基性タンパク質(ヒストン)がありません。 通常、細菌細胞には 1 つの染色体が含まれており、これは環状に閉じた DNA 分子で表されます。 分裂が中断されている場合は、4 本以上の染色体が含まれている可能性があります。 核様体は、DNA 固有の方法 (フォイゲン法またはロマノフスキー ギムザ法) を使用して染色した後、光学顕微鏡で検出されます。 細菌の極薄切片の電子回折パターンでは、核様体は、特定の領域で細胞質膜または染色体の複製に関与するメソソームに結合した DNA の原線維状の糸状構造を備えた明るいゾーンとして見えます。
1 本の染色体によって表される核様体に加えて、細菌細胞には染色体外遺伝因子が含まれています。 プラスミド, これは共有結合で閉じた DNA の環です。
カプセル - 厚さ0.2ミクロンを超える粘液構造で、細菌の細胞壁としっかりと結合しており、明確に定義された外部境界を持っています。 カプセルは、病理学的物質からの痕跡のスミアで見ることができます。 純粋な細菌培養物では、莢膜が形成される頻度は低くなります。 これは、カプセル物質の負のコントラストを生み出す特別な Burri-Gins 染色法を使用して検出されます。
通常、莢膜は多糖類 (エキソ多糖類) で構成されており、例えば炭疽菌ではポリペプチドで構成されることもあります。 カプセルは親水性で、細菌の貪食を防ぎます。
多くの細菌が発生します マイクロカプセル -厚さ0.2ミクロン未満の粘液形成。電子顕微鏡でのみ検出可能。 カプセルとは区別する必要があります スライム -明確な外部境界を持たないムコイド外多糖。 ムコイド外多糖は、嚢胞性線維症患者の喀痰中によく見られる緑膿菌のムコイド株の特徴です。 細菌のエキソ多糖は接着 (基質への付着) に関与しており、細菌外多糖とも呼ばれます。 グリコカリックス。細菌による細胞外多糖類の合成に加えて、その形成には別のメカニズムがあります。それは、二糖類に対する細胞外細菌酵素の作用によるものです。 その結果、デキストランとレバンが形成されます。 カプセルと粘液は、親水性であるため、水とよく結合し、マクロ微生物やバクテリオファージの保護因子の作用を防ぐため、細菌を損傷や乾燥から守ります。
鞭毛細菌は細菌細胞の移動性を決定します。 鞭毛は細胞質膜に由来する細い糸状で、細胞自体よりも長いです(図3)。 鞭毛の太さは12~20nm、長さは3~12μmです。 細菌の種類によって鞭毛の数は異なります。 (モノリッチ)コレラビブリオ菌には、細菌の周囲に沿って伸びる最大数十、数百の鞭毛があります。 (ペリトリチ)大腸菌、プロテウスなど ロフォトリクス細胞の一端に鞭毛の束を持っています。 両親媒性疾患細胞の両端に1本の鞭毛または鞭毛の束を持っています。 鞭毛は特別なディスクによって細胞質膜と細胞壁に付着しています。 鞭毛はタンパク質 - フラジェリン(から naT.鞭毛 -鞭毛)、抗原特異性を持っています。 フラジェリンのサブユニットはらせん状にねじれています。 鞭毛は、重金属でコーティングされた製剤の電子顕微鏡を使用して検出するか、鞭毛の厚さを増加させるさまざまな物質のエッチングと吸着に基づく特別な方法で処理した後(たとえば、銀メッキ後)の光学顕微鏡で検出します。

米。 3. 大腸菌。 電子回折パターン (V.S. Tyurin による作成)。 1 - 鞭毛、2 - 絨毛、3 - F-線毛。
絨毛、または線毛 (線毛)、 - 糸状の構造(図 3)、鞭毛よりも薄くて短い(3 ~ 10 nm x 0.3 ~ 10 μm)。 線毛は細胞表面から伸びており、タンパク質線毛で構成されています。 それらは抗原活性を持っています。 線毛の中には以下のものがあります: 接着を担当する線毛、つまり 影響を受けた細胞への細菌の付着(タイプ 1 線毛、または一般的なタイプ - 共通線毛)、飲酒し、栄養、水と塩の代謝を担当します。 性的 (飲んだ)、または共役線毛 (タイプ 2 線毛)。 一般的なタイプの線毛は多数あり、1細胞あたり数百個です。 性線毛は、伝達可能なプラスミドを含むいわゆる「男性」ドナー細胞によって形成されます (F、R、Col)。通常、1 つのセルに 1 ~ 3 個あります。 性線毛の際立った特徴は、性線毛に集中的に吸着される特別な「男性」球状バクテリオファージとの相互作用です。
論争 - 休止中のファーミキュート細菌の特異な形態、すなわち グラム陽性タイプの細胞壁構造を持つ細菌。
胞子は細菌の存在にとって不利な条件(乾燥、栄養不足など)下で形成されます。 この場合、一つの細菌の中に一つの胞子が形成されます。 胞子の形成は種の保存に寄与するものであり、真菌の場合のように繁殖の手段ではありません。
胞子のサイズが細胞の直径を超えない、胞子形成好気性細菌と呼ばれることもあります。 桿菌。胞子のサイズが細胞の直径を超え、紡錘形の形状をとる芽胞形成性嫌気性細菌を胞子形成性嫌気性細菌と呼びます。 クロストリジウム症(緯度。 クロストリジウム-スピンドル)。
プロセス 胞子形成(胞子形成) は一連の段階を経て、細胞質の一部と染色体が分離され、細胞質膜に囲まれます。 胞子が形成され、次に多層の透過性の低い殻が形成されます。 胞子形成には、胞子が集中的に消費され、その後、ジピコリン酸とカルシウムイオンの胞子殻が形成されます。 すべての構造が形成された後、胞子は耐熱性を獲得しますが、これはジピコリン酸カルシウムの存在に関連しています。 胞子形成、細胞内での胞子の形状と位置 (栄養) は細菌の種の特性であり、細菌を互いに区別することができます。 胞子の形状は楕円形、球形であり、細胞内の位置は末端、つまり末端です。 棒の端(破傷風の原因物質)、サブターミナル - 棒の端に近い(ボツリヌス症、ガス壊疽の病原体)、および中央(炭疽菌)。
細菌は、現在地球上に存在する最も古い生物群です。 最初の細菌はおそらく 35 億年以上前に出現し、ほぼ 10 億年間、地球上で唯一の生物でした。 これらは生きた自然の最初の代表者であったため、彼らの体は原始的な構造を持っていました。
時間が経つにつれて、その構造はより複雑になりましたが、今日に至るまで、細菌は最も原始的な単細胞生物であると考えられています。 興味深いことに、一部の細菌は古代の祖先の原始的な特徴をまだ保持しています。 これは、高温の硫黄泉や貯水池の底の無酸素泥に生息する細菌で観察されます。
ほとんどの細菌は無色です。 紫や緑のものはほんのわずかです。 しかし、多くの細菌のコロニーは明るい色をしています。これは、環境への着色物質の放出または細胞の色素沈着によって引き起こされます。
細菌の世界の発見者は、17 世紀のオランダの博物学者、アントニー レーウェンフックであり、彼は物体を 160 ~ 270 倍に拡大する完璧な拡大顕微鏡を初めて作成しました。
細菌は原核生物として分類され、別の界である細菌に分類されます。
体型
細菌は多数の多様な生物です。 形状はさまざまです。
| 細菌の名前 | 細菌の形状 | 細菌画像 |
| 球菌 | ボール状 | |
| バチルス |  | 棒状 |
| ビブリオ | 勾玉 | |
| スピリラム |  | 螺旋 |
| 連鎖球菌 |  | 球菌の連鎖 |
| ブドウ球菌 |  | 球菌の塊 |
| 双球菌 | 1つの粘膜カプセルの中に2つの丸い細菌が封入されている |
交通手段
細菌の中には、移動するものと移動しないものがあります。 運動体は、波状の収縮によって、またはフラジェリンと呼ばれる特殊なタンパク質からなる鞭毛(ねじれたらせん状の糸)の助けを借りて動きます。 鞭毛は 1 つ以上存在する場合があります。 いくつかの細菌では、それらは細胞の一端に位置し、他の細菌では二端または表面全体に位置します。

しかし、運動は鞭毛を持たない他の多くの細菌にも固有のものです。 したがって、粘液で外側を覆われた細菌は滑走運動が可能です。
鞭毛を持たない水生細菌や土壌細菌の中には、細胞質内にガス空胞を持つものがあります。 細胞には 40 ~ 60 個の液胞が存在する場合があります。 それらのそれぞれにはガス(おそらく窒素)が充填されています。 液胞内のガスの量を調節することにより、水生細菌は水柱に沈んだり、水面に浮上したりすることができ、土壌細菌は土壌の毛細管内を移動することができます。
生息地
組織の単純さと気取らないため、細菌は自然界に広く普及しています。 細菌はどこにでも存在します。たとえ最も純粋な湧き水の一滴でも、土の粒の中、空気中、岩の上、極地の雪の中、砂漠の砂の中、海底、深いところから抽出された油の中、さらには地下水の中にも存在します。温度約80℃の温泉水。 植物、果物、さまざまな動物、そして人間の腸内に生息しており、 口腔、手足に、体の表面に。
細菌は最も小さく、最も数の多い生き物です。 サイズが小さいため、亀裂、隙間、または毛穴に簡単に浸透します。 非常に丈夫で、さまざまな生活条件に適応します。 生存能力を失うことなく、乾燥、極寒、90℃までの加熱に耐えます。
地球上でバクテリアが存在しない場所は事実上ありませんが、その量はさまざまです。 細菌の生存条件はさまざまです。 それらの中には、大気中の酸素を必要とするものもあれば、それを必要とせず、酸素のない環境でも生きられるものもあります。
空気中: バクテリアは大気上層 30 km まで上昇します。 もっと。
特に土壌中には多く存在します。 1 g の土壌には数億の細菌が含まれている可能性があります。
水中: 開いた貯水池の水の表層。 有益な水生バクテリアが有機残留物を石化します。
生物の場合:病原性細菌は外部環境から体内に侵入しますが、病気を引き起こすのは好ましい条件下に限られます。 共生生物は消化器官に生息し、食物の分解と吸収、ビタミンの合成を助けます。
外部構造
細菌の細胞は、特別な高密度の殻である細胞壁で覆われており、保護機能と支持機能を果たし、細菌に永続的な特徴的な形状を与えます。 細菌の細胞壁は植物の細胞壁に似ています。 それは透過性です。それを通して、栄養素は自由に細胞に入り、代謝産物は環境に出ます。 多くの場合、細菌は細胞壁の上に粘液の追加の保護層、つまりカプセルを生成します。 カプセルの厚さは細胞自体の直径よりも何倍も大きくなる場合もありますが、非常に小さくなる場合もあります。 カプセルは細胞の必須部分ではなく、細菌が置かれる条件に応じて形成されます。 細菌を乾燥から守ります。
一部の細菌の表面には、長い鞭毛 (1 つ、2 つ、または多数) または短くて薄い絨毛があります。 鞭毛の長さは細菌の体の大きさよりも何倍も大きくなることがあります。 細菌は鞭毛と絨毛の助けを借りて移動します。

内部構造
細菌細胞の内部には、高密度で不動の細胞質があります。 層状構造をしており、液胞がないため、細胞質そのものの中にさまざまなタンパク質(酵素)や予備栄養素が存在します。 細菌の細胞には核がありません。 遺伝情報を伝える物質は細胞の中心部分に集中しています。 細菌、核酸、DNA。 しかし、この物質は核にはなりません。
細菌細胞の内部組織は複雑で、それぞれに特有の特徴があります。 細胞質は細胞膜によって細胞壁から分離されています。 細胞質には、主要な物質、つまりマトリックス、リボソーム、およびさまざまな機能を実行する少数の膜構造(ミトコンドリア、小胞体、ゴルジ装置の類似体)があります。 細菌細胞の細胞質には、さまざまな形状やサイズの顆粒が含まれていることがよくあります。 顆粒は、エネルギー源および炭素源として機能する化合物で構成されている場合があります。 脂肪の液滴は細菌細胞にも見られます。
細胞の中心部には、核物質であるDNAが局在しており、膜によって細胞質と区切られていません。 これは核の類似体である核様体です。 核様体には、膜、核小体、または染色体のセットがありません。
食事方法
細菌にはさまざまな摂食方法があります。 その中には独立栄養生物と従属栄養生物があります。 独立栄養生物は、栄養となる有機物質を独立して生産できる生物です。
植物は窒素を必要としますが、植物自体は空気中の窒素を吸収できません。 一部の細菌は、空気中の窒素分子と他の分子を結合させ、植物が利用できる物質を生成します。
これらの細菌は若い根の細胞に定着し、結節と呼ばれる根の肥厚を引き起こします。 このような根粒は、マメ科の植物や他のいくつかの植物の根に形成されます。
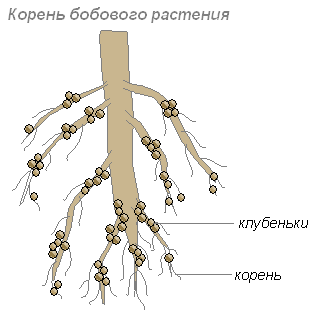
根は細菌に炭水化物を提供し、細菌は根に植物が吸収できる窒素含有物質を提供します。 彼らの同棲は相互に利益をもたらします。
植物の根からは多くの分泌物が分泌されます 有機物細菌の餌となる(糖類、アミノ酸など)。 そのため、根の周囲の土壌層には特に多くの細菌が定着します。 これらの細菌は、植物の死骸を植物が利用できる物質に変換します。 この土壌の層は根圏と呼ばれます。
根粒細菌の根組織への侵入については、いくつかの仮説があります。
- 表皮および皮質組織への損傷による。
- 根毛を通して。
- 若い細胞膜のみを通過します。
- ペクチン分解酵素を生成する仲間バクテリアのおかげ。
- 植物の根の分泌物中に常に存在するトリプトファンからの B-インドール酢酸の合成が刺激されるためです。
根粒細菌を根組織に導入するプロセスは、次の 2 つの段階で構成されます。
- 根毛の感染。
- 結節形成のプロセス。
ほとんどの場合、侵入した細胞は活発に増殖し、いわゆる感染糸を形成し、その糸の形で植物組織内に移動します。 感染糸から出現した結節細菌は宿主組織内で増殖し続けます。

急速に増殖する根粒細菌の細胞で満たされた植物細胞は、急速に分裂を始めます。 若い結節とマメ科植物の根との接続は、維管束線維束のおかげで行われます。 機能している期間中、結節は通常密集しています。 最適な活動が起こるまでに、小結節はピンク色になります (レグヘモグロビン色素のおかげです)。 レグヘモグロビンを含む細菌だけが窒素を固定することができます。
根粒菌は土壌 1 ヘクタールあたり数十、数百キログラムの窒素肥料を生成します。
代謝
細菌はそれぞれ代謝が異なります。 酸素の参加によって起こる人もいれば、酸素なしで起こる人もいます。
ほとんどの細菌は既製の有機物質を餌とします。 無機物から有機物を作り出すことができるのは、そのうちの少数のバクテリア (青緑色バクテリア、またはシアノバクテリア) だけです。 それらは地球の大気中の酸素の蓄積において重要な役割を果たしました。
細菌は外部から物質を吸収し、その分子を細かく引き裂き、その部分から殻を組み立てて内容物を補充し(これが細菌の増殖方法です)、不要な分子を捨てます。 細菌の殻と膜は、細菌がのみを吸収できるようにします。 必要な物質.

細菌の殻や膜が完全に不透過性であれば、物質は細胞内に侵入しません。 もしそれらがすべての物質に対して透過性であれば、細胞の内容物は培地、つまり細菌が生息する溶液と混合することになります。 細菌が生きていくためには、必要な物質は通過させるが、不要な物質は通過させない殻が必要です。
細菌は近くにある栄養素を吸収します。 次は何が起こる? 独立して移動できる場合(鞭毛を動かすか、粘液を押し戻すことによって)、必要な物質を見つけるまで移動します。

移動できない場合は、拡散(ある物質の分子が別の物質の分子の茂みに浸透する能力)によって必要な分子がそこに運ばれるまで待ちます。
細菌は、他の微生物群とともに膨大な化学的仕事を行います。 さまざまな化合物を変換することで、生命に必要なエネルギーや栄養素を摂取しています。 細菌の代謝プロセス、エネルギーの獲得方法、体の物質を構築するための材料の必要性は多様です。
他の細菌は、無機化合物を犠牲にして、体内の有機物質の合成に必要な炭素の需要をすべて満たします。 それらは独立栄養生物と呼ばれます。 独立栄養細菌は、無機物から有機物を合成することができます。 その中には次のようなものがあります。
化学合成
放射エネルギーの利用は最も重要ですが、二酸化炭素と水から有機物を生成する唯一の方法ではありません。 細菌は、そのような合成のエネルギー源として太陽光ではなくエネルギーを使用することが知られている。 化学結合、特定の無機化合物(硫化水素、硫黄、アンモニア、水素、硝酸、第一鉄化合物、マンガン)の酸化中に生物の細胞内で発生します。 彼らは、この化学エネルギーを使用して形成された有機物を使用して、体の細胞を構築します。 したがって、このプロセスは化学合成と呼ばれます。
化学合成微生物の最も重要なグループは硝化細菌です。 これらの細菌は土壌に生息し、有機残留物の分解中に生成されるアンモニアを酸化して硝酸にします。 後者は土壌の鉱物化合物と反応して硝酸の塩に変わります。 このプロセスは 2 つのフェーズで行われます。
鉄バクテリアは第一鉄を酸化鉄に変換します。 結果として生じる水酸化鉄は沈殿し、いわゆる湿原鉄鉱石を形成します。
一部の微生物は水素分子の酸化によって存在し、それによって栄養の独立栄養法を提供します。
水素バクテリアの特徴は、有機化合物が供給され、水素が存在しない場合に従属栄養的な生活様式に切り替える能力です。
したがって、化学独立栄養生物は、無機物質から必要な有機化合物を独立して合成し、従属栄養生物のように他の生物から既製のものを摂取しないため、典型的な独立栄養生物です。 化学独立栄養細菌は、エネルギー源としての光から完全に独立しているという点で、光合成植物とは異なります。
細菌の光合成
特定の色素であるバクテリオクロロフィルを含む一部の色素含有硫黄バクテリア(紫、緑)は、バクテリオクロロフィルを吸収することができます。 太陽光エネルギー、その助けを借りて、体内の硫化水素が分割され、水素原子を放出して、対応する化合物を復元します。 このプロセスは光合成と多くの共通点がありますが、紫色および緑色の細菌では水素供与体が硫化水素(場合によってはカルボン酸)であり、緑色植物ではそれが水であるという点のみが異なります。 どちらも吸収した太陽光線のエネルギーにより水素の分離・移動が行われます。
酸素を放出せずに起こるこの細菌の光合成は、光還元と呼ばれます。 二酸化炭素の光還元は、水からではなく硫化水素からの水素の移動に関連しています。
6СО 2 +12Н 2 S+hv → С6Н 12 О 6 +12S=6Н 2 О
地球規模での化学合成と細菌の光合成の生物学的重要性は比較的小さい。 自然界における硫黄循環のプロセスにおいて重要な役割を果たすのは化学合成細菌だけです。 吸収された 緑の植物硫酸塩の形で硫黄は還元され、タンパク質分子の一部になります。 さらに、植物や動物の死骸が腐敗細菌によって破壊されると、硫黄が硫化水素の形で放出され、硫化水素が硫黄細菌によって酸化されて硫黄(または硫酸)が遊離され、植物が利用できる亜硫酸塩が土壌中に形成されます。 化学独立栄養細菌と光合成独立栄養細菌は、窒素と硫黄のサイクルに不可欠です。
胞子形成
胞子は細菌の細胞内で形成されます。 胞子形成の過程で、細菌細胞は多くの生化学的プロセスを経ます。 その中の自由水の量が減少し、酵素活性が低下します。 これにより、好ましくない環境条件(高温、高塩濃度、乾燥など)に対する胞子の耐性が確保されます。 胞子形成は、少数の細菌のグループのみに特徴的です。

紛争は必須の段階ではない ライフサイクル細菌。 胞子形成は、栄養素の欠乏または代謝産物の蓄積によってのみ始まります。 胞子の形をした細菌は、長期間休眠状態に留まることがあります。 細菌の胞子は、長時間の煮沸や非常に長時間の冷凍に耐えることができます。 好ましい条件が発生すると、胞子は発芽し、生存可能になります。 細菌の胞子は、不利な条件で生き残るための適応です。
再生
細菌は1つの細胞を2つに分裂させることで繁殖します。 細菌は一定の大きさに達すると、2 つの同一の細菌に分裂します。 その後、それぞれが摂食を開始し、成長し、分裂します。

細胞伸長後、横隔壁が徐々に形成され、その後娘細胞が分離します。 多くの細菌では、特定の条件下では、分裂後、細胞は特徴的なグループに接続されたままになります。 この場合、分割面の方向や分割数によって異なる形状が生じます。 出芽による生殖は細菌では例外的に起こります。
良好な条件下では、多くの細菌の細胞分裂は 20 ~ 30 分ごとに起こります。 このような急速な繁殖により、5 日以内に 1 つの細菌の子孫が、すべての海と海洋を埋めることができる塊を形成する可能性があります。 単純に計算すると、1 日に 72 世代 (720,000,000,000,000,000,000 個の細胞) が形成されることがわかります。 重量に換算すると4720トン。 しかし、ほとんどの細菌は環境にさらされるとすぐに死滅するため、自然界ではこのようなことは起こりません。 日光、乾燥中、食物の不足、種間の闘争の結果として65〜100℃に加熱するなど。
細菌 (1) は十分な食物を吸収すると、サイズが大きくなり (2)、生殖 (細胞分裂) の準備を開始します。 その DNA (細菌では DNA 分子は環状に閉じられています) は 2 倍になります (細菌はこの分子のコピーを生成します)。 両方の DNA 分子 (3、4) は細菌の壁に付着していることに気づき、細菌が伸長するにつれて離れていきます (5、6)。 まずヌクレオチドが分裂し、次に細胞質が分裂します。

2 つの DNA 分子が分岐した後、細菌にくびれが現れ、細菌の体が徐々に 2 つの部分に分割され、それぞれの部分に DNA 分子が含まれます (7)。
(枯草菌では) 2 つの細菌がくっつき、それらの間に橋が形成されることが起こります (1、2)。

ジャンパーは DNA をある細菌から別の細菌に輸送します (3)。 1 つの細菌に入ると、DNA 分子が絡み合い、いくつかの場所でくっつき (4)、その後セクションを交換します (5)。
自然界における細菌の役割
ジャイア
細菌は、自然界の物質の一般的な循環において最も重要なつながりです。 植物は土壌中の二酸化炭素、水、無機塩から複雑な有機物質を生成します。 これらの物質は死んだ菌類、植物、動物の死骸とともに土壌に戻ります。 細菌は複雑な物質を単純な物質に分解し、それを植物が利用します。
細菌は、植物や動物の死骸、生物の排泄物、さまざまな廃棄物の複雑な有機物質を破壊します。 これらの有機物質を餌とする腐生細菌は、それらを腐植に変えます。 これらは私たちの惑星の一種の秩序です。 このように、細菌は自然界の物質循環に積極的に参加しています。
土壌形成
細菌はほぼあらゆる場所に分布し、膨大な数で発生するため、自然界で起こるさまざまなプロセスを大きく決定します。 秋には、木や低木の葉が落ち、地上の草の芽は枯れ、古い枝は落ち、時には古い木の幹が倒れます。 これらすべてが徐々に腐植に変わります。 1cm3で。 森林土壌の表層には、数億種の腐生土壌細菌が含まれています。 これらのバクテリアは腐植をさまざまな物質に変換します。 ミネラル植物の根によって土壌から吸収されます。
一部の土壌バクテリアは空気から窒素を吸収し、それを重要なプロセスに使用することができます。 これらの窒素固定細菌は独立して生息するか、マメ科植物の根に定着します。 マメ科植物の根に侵入したこれらの細菌は、根細胞の成長とその根粒の形成を引き起こします。
これらの細菌は植物が利用する窒素化合物を生成します。 細菌は植物から炭水化物と無機塩を入手します。 したがって、マメ科植物と根粒菌の間には密接な関係があり、一方の生物と他方の生物の両方にとって有益です。 この現象を共生といいます。
マメ科植物は根粒菌との共生のおかげで土壌を窒素で豊かにし、収量の増加に役立ちます。
自然界における分布
微生物はどこにでも存在します。 唯一の例外は、活火山のクレーターと爆発した原子爆弾の震源地の小さな地域です。 南極の低温も、間欠泉の沸騰する流れも、塩だまりの飽和食塩水も、山頂の強い日射も、原子炉の過酷な放射線も、微生物相の存在と発達を妨げません。 すべての生き物は常に微生物と相互作用しており、微生物の保管庫であるだけでなく、配布者でもあることがよくあります。 微生物は私たちの地球の在来種であり、最も驚くべき自然の基質を積極的に探索しています。
土壌微生物相
土壌中の細菌の数は非常に多く、1グラムあたり数億、数十億の細菌が存在します。 それらは水中や空気中よりも土壌中にはるかに多く存在します。 土壌中の細菌の総数が変化します。 細菌の数は土壌の種類、状態、地層の深さによって異なります。
土壌粒子の表面では、微生物は小さな微小コロニー (それぞれ 20 ~ 100 個の細胞) に存在します。 それらは多くの場合、有機物の塊の厚さ、生きている植物の根や枯れかけている植物の根、細い毛細血管、塊の内部に発生します。
土壌微生物相は非常に多様です。 ここには、腐敗細菌、硝化細菌、窒素固定細菌、硫黄細菌などのさまざまな生理学的細菌群があり、その中には好気性菌と嫌気性菌、胞子型と非胞子型が存在します。 微生物相は土壌形成の要因の 1 つです。
土壌中の微生物の発達領域は、生きている植物の根に隣接するゾーンです。 それは根圏と呼ばれ、そこに含まれる微生物の総体は根圏微生物叢と呼ばれます。
貯水池の微生物叢
水は微生物が多く生息する自然環境です。 それらの大部分は土壌から水に入ります。 水中のバクテリアの数と栄養素の存在を決定する要素。 最もきれいな水は自噴井戸や泉からのものです。 開いた貯水池や川には細菌が非常に豊富です。 細菌の数が最も多いのは、海岸に近い水の表層です。 岸から遠ざかり、深くなるとバクテリアの数は減少します。
きれいな水には 1 ml あたり 100 ~ 200 個の細菌が含まれており、汚染された水には 10 万~30 万個以上の細菌が含まれています。 底泥、特に表層にはバクテリアが多く存在しており、バクテリアが膜を形成しています。 この膜には硫黄バクテリアや鉄バクテリアが多く含まれており、硫化水素を酸化して硫酸にし、魚の死滅を防ぎます。 シルト中には胞子を有する形態がより多く存在するが、水中では胞子を有さない形態が優勢である。
種構成の点では、水の微生物相は土壌の微生物相と似ていますが、特定の形態もあります。 微生物は水中に入るさまざまな廃棄物を破壊することによって、いわゆる水の生物学的浄化を徐々に実行します。
空気微生物叢
空気の微生物相は、土壌や水の微生物相に比べて数が少ないです。 細菌は塵とともに空気中に舞い上がり、しばらくそこに留まり、その後地表に定着し、栄養不足や紫外線の影響で死んでしまいます。 空気中の微生物の数は、地理的ゾーン、地形、季節、粉塵汚染などによって異なります。粉塵の各点は微生物の媒介者です。 ほとんどの細菌は産業上の空気中に存在します。 田舎のほうが空気がきれいです。 最もきれいな空気は森林、山、雪の多い地域の上にあります。 空気の上層には微生物がほとんど含まれていません。 空気中の微生物叢には、色素を帯びた胞子を持つ細菌が多く含まれており、他の細菌よりも紫外線に対して耐性があります。
人体の微生物叢
人間の体は、完全に健康な体であっても、常に微生物叢の保菌者です。 人体が空気や土壌と接触すると、病原性微生物(破傷風菌、ガス壊疽など)を含むさまざまな微生物が衣服や皮膚に定着します。 露出した部分が最も汚染されやすいです。 人体。 手には大腸菌やブドウ球菌が見られます。 口腔内には100種類以上の微生物が存在します。 温度、湿度、栄養素が残留している口は、微生物の発育に最適な環境です。
胃は酸性反応を示すため、胃内の微生物の大部分は死滅します。 小腸から始まり、反応はアルカリ性になります。 微生物にとって有利です。 大腸内の微生物叢は非常に多様です。 成人一人当たり、毎日約 180 億個の細菌が排泄物として排泄されます。 地球上では人間よりも個人の方が多いのです。
外部環境とつながっていない内臓(脳、心臓、肝臓、膀胱など)には通常、微生物が存在しません。 微生物は病気のときにのみこれらの臓器に入ります。
物質循環における細菌
一般に微生物、特に細菌は、植物や動物がまったくアクセスできない化学変化を実行し、地球上の生物学的に重要な物質の循環において大きな役割を果たしています。 元素のサイクルのさまざまな段階は、さまざまな種類の生物によって実行されます。 生物の個々のグループの存在は、他のグループによって実行される元素の化学変化に依存します。
窒素循環
窒素化合物の循環変換は、さまざまな栄養ニーズを持つ生物圏の生物に必要な形態の窒素を供給する上で主要な役割を果たします。 総窒素固定の 90% 以上は、特定の細菌の代謝活動によるものです。
炭素循環
分子状酸素の還元を伴う有機炭素の二酸化炭素への生物学的変換には、さまざまな微生物の共同代謝活動が必要です。 多くの好気性細菌は有機物の完全な酸化を実行します。 好気的条件下では、有機化合物は発酵によって最初に分解され、無機水素受容体(硝酸塩、硫酸塩、またはCO 2 )が存在する場合、発酵の最終有機生成物は嫌気呼吸によってさらに酸化されます。
硫黄サイクル
硫黄は、主に可溶性硫酸塩または還元された有機硫黄化合物の形で生物に利用されます。
鉄サイクル
一部の淡水域には高濃度の還元鉄塩が含まれています。 そのような場所では、還元鉄を酸化する鉄バクテリアという特定の細菌叢が発達します。 彼らは湿原鉄鉱石と鉄塩が豊富な水源の形成に関与しています。
細菌は最も古い生物であり、始生代の約 35 億年前に出現しました。 約25億年間、彼らは地球を支配して生物圏を形成し、酸素大気の形成に参加しました。
細菌は、(ウイルスを除いて) 最も単純な構造の生物の 1 つです。 彼らは地球上に出現した最初の生物であると考えられています。
細菌細胞の構成要素は 2 つのタイプに分類されます。
- 基本構造(細胞壁、細胞質膜とその誘導体、細胞質とリボソームおよびさまざまな封入体、核様体);
- 仮設構造物(莢膜、粘膜、鞭毛、絨毛、内生胞子、細菌の生活環の特定の段階でのみ形成されます)。
基本的な構造。
細胞壁細胞膜の外側に位置します。 細胞質膜は細胞壁の一部ではありません。 細胞壁の機能:
浸透圧ショックやその他の有害な要因から細菌を保護します。
細菌の形状の決定;
細菌の代謝への参加。
細胞壁には細孔が浸透しており、そこを通って細菌の外毒素が輸送されます。 細胞壁の厚さは10〜100nmです。 細菌の細胞壁の主成分は、 ペプチドグリカンまたは ムレイン、グリコシド結合によって接続された交互の N-アセチル-N-グルコサミンと N-アセチルムラミン酸残基から構成されます。
1884 年、H. グラムは、リンドウ バイオレット、ヨウ素、エチル アルコール、フクシンを使用して細菌を染色する方法を提案しました。 すべての細菌は、グラム染色に応じて、グラム陽性菌とグラム陰性菌の 2 つのグループに分類されます。 グラム陽性菌の細胞壁細胞膜にしっかりとフィットし、その厚さは20〜100 nmです。 これには、テイコ酸(グリセロールまたはリビトールのポリマー)のほか、少量の多糖類、タンパク質、脂質が含まれています。 グラム陰性菌の細胞壁多層で、その厚さは14〜17 nmです。 内層(ペプチドグリカン)は薄い連続ネットワークを形成します。 外層はリン脂質、リポタンパク質、タンパク質で構成されています。 外膜タンパク質はペプチドグリカン層にしっかりと結合しています。
特定の条件下では、細菌は細胞壁成分を完全または部分的に合成する能力を失い、その結果、プロトプラスト、スフェロプラスト、および L 型細菌が形成されます。 スフェロプラスト細胞壁が部分的に破壊された細菌です。 それらはグラム陰性菌で観察されます。 プロトプラスト- これらは細胞壁が完全に欠如した形態です。 それらはグラム陽性菌によって形成されます。 L字型細菌は、細胞壁ペプチドグリカンを合成する能力を部分的または完全に失った細菌の突然変異体(細胞壁に欠陥のある細菌)です。 彼らの名前は、1935 年に設立されたイギリスのリスター研究所の名前に由来しています。
細胞質膜 (CPM) とその誘導体。細胞質膜 (原形質膜) は、細胞壁から細胞質を分離する細菌細胞の半透過性リポタンパク質構造です。 それは細胞の乾燥質量の 8 ~ 15% を占めます。 その破壊は細胞死につながります。 電子顕微鏡検査により、その三層構造が明らかになりました。 細胞質膜はタンパク質 (50 ~ 75%) と脂質 (15 ~ 20%) の複合体です。 脂質の大部分はリン脂質に代表されます。 さらに、少量の炭水化物が膜内に検出されました。
バクテリア CPM は次の機能を実行します。
バリア機能 (分子の「ふるい」)。
エネルギー;
特別なキャリア(トランスロカーゼまたはパーミアーゼ)を使用した、さまざまな有機および無機の分子およびイオンの選択的移動。
複製とその後の染色体分裂。
細胞の成長中、細胞質膜は、と呼ばれる多数の陥入 (陥入) を形成します。 メソソーム.
細胞質 -これは細菌細胞の内容物であり、細胞質膜で囲まれています。 それはサイトゾルと構造要素から構成されます。
サイトゾル- 可溶性 RNA 成分、酵素、代謝産物を含む均一な画分。
構造要素- これらはリボソーム、細胞質内膜、封入体および核様体です。
リボソーム- タンパク質生合成を実行する細胞小器官。 それらはタンパク質とRNAから構成されます。 直径15~20nmの顆粒です。 1 つの細菌細胞には 5,000 ~ 50,000 のリボソームが含まれています。 リボソームはタンパク質合成の部位です。
原核生物の細胞質には、細胞の予備物質を表すさまざまな封入体が見られます。 多糖類のうち、グリコーゲン、デンプン、およびデンプン様物質である顆粒膜は細胞内に沈着します。 ポリリン酸塩は、と呼ばれる顆粒に含まれています。 ボリューティニック、 または 異染性の、 穀類。
核様体原核生物の核です。 これは環状に閉じた 1 本の二本鎖 DNA 鎖で構成されており、細菌の染色体と考えられています。 核様体には核膜がありません。
核様体に加えて、染色体外の遺伝要素が細菌細胞で見つかりました。 プラスミド、自律複製可能な小さな環状 DNA 分子です。 プラスミドの役割は、特定の生存条件において細胞に利点を与える追加の特性をコードすることです。 最も一般的なプラスミドは、細菌の抗生物質耐性の兆候 (R プラスミド)、エンテロトキシン (Ent プラスミド) または溶血素 (Hly プラスミド) の合成の兆候を決定するプラスミドです。
に 仮設構造物莢膜、鞭毛、線毛、細菌の内生胞子が含まれます。
カプセル -これは細菌の細胞壁上の粘膜層です。 カプセル物質は多糖類の糸で構成されています。 カプセルは細胞質膜の外表面で合成され、特定の領域の細胞壁表面に放出されます。
カプセルの機能:
細菌の毒性、抗原特異性、免疫原性を決定する莢膜抗原の位置。
機械的損傷、乾燥、有毒物質、ファージによる感染、マクロ微生物の保護因子の作用からの細胞の保護。
細胞が基質に付着する能力。
鞭毛 –これらは細菌が活動する器官です。 鞭毛は重要な構造物ではないため、増殖条件に応じて細菌内に存在する場合と存在しない場合があります。 鞭毛の数とその位置は細菌によって異なります。 これに応じて、次の鞭毛細菌のグループが区別されます。
- モノリッチ– 1 つの極性の鞭毛を持つ細菌。
- アンフィトリクス– 極性を持って配置された 2 本の鞭毛を持つ細菌、または両端に鞭毛の束を持つ細菌。
- ロフォトリクス– 細胞の一端に鞭毛の束を持つ細菌。
- 周縁性- 細胞の側面または表面全体に多数の鞭毛を持つ細菌。
鞭毛の化学組成はタンパク質で表されます フラジェリン.
細菌細胞の表面構造には次のものもあります。 絨毛そして 飲んだ。 これらの構造は、基質(絨毛、一般線毛)への細胞の吸着および遺伝物質の伝達プロセス(性線毛)に関与しています。 それらは特定の疎水性タンパク質によって形成されます ピリン。
一部の細菌では、特定の条件下で休眠形態が形成され、不利な条件下でも細胞が長期間生存できるようになります。 内生胞子。 有害な環境要因に対して耐性があります。
細胞内の胞子の位置:
中枢性(炭疽菌の原因物質)。
サブターミナル - 末端に近い(ボツリヌス症の原因物質)。
ターミナル - スティックの端(破傷風の原因物質)。
生きた自然の 5 つの王国に加えて、さらに 2 つの超王国、原核生物と真核生物があります。 したがって、細菌の系統的な位置を考慮すると、次のようになります。
これらの生物はなぜ別の分類群として分類されるのでしょうか? 重要なのは、細菌細胞は、その生命活動や他の生き物や人間との相互作用に痕跡を残す特定の特徴の存在によって特徴付けられるということです。
細菌の発見

リボソームは、細胞質内に多数散在する小さな構造です。 それらの性質は RNA 分子によって表されます。 これらの顆粒は、特定の種類の細菌の関係の程度と系統的な位置を決定できる材料です。 それらの機能はタンパク質分子の集合です。
カプセル
細菌細胞は保護粘膜の存在を特徴とし、その組成は多糖類またはポリペプチドによって決定されます。 このような構造はカプセルと呼ばれます。 マイクロカプセルとマクロカプセルがあります。 この構造はすべての種で形成されるわけではありませんが、大部分の種で形成される、つまり必須ではありません。
カプセルは細菌細胞を何から保護しますか? 細菌が病原性の場合、宿主抗体による食作用による。 他のタイプについて言えば、乾燥や有害物質への曝露によるものもあります。
粘液と封入物
細菌の任意の構造も。 粘液、または糖衣は、化学的にはムコイド多糖に基づいています。 それは細胞内でも、外部酵素によっても形成されます。 水によく溶けます。 目的: 基材への細菌の付着 - 接着。
封入体は、細胞質内のさまざまな化学的性質の微粒子です。 これらは、タンパク質、アミノ酸、核酸、または多糖類であってもよい。

動きのオルガノイド
細菌の細胞の特徴はその動きにも表れます。 この目的のために、鞭毛がさまざまな数(細胞あたり 1 個から数百個まで)存在します。 各鞭毛の基礎はタンパク質であるフラジェリンです。 弾性収縮と左右へのリズミカルな動きのおかげで、細菌は空間を移動できます。 鞭毛は細胞膜に付着しています。 場所によっても異なる場合があります 他の種類.
飲んだ
鞭毛よりもさらに細かい構造は、次のことに関与します。
- 基材への付着。
- 水と塩の栄養。
- 有性生殖。
それらはタンパク質ピリンで構成されており、その数は細胞あたり数百に達することがあります。
植物細胞との類似点
細菌との間違いのない類似点は、細胞壁の存在です。 ただし、植物では間違いなく存在しますが、細菌ではすべての種に存在するわけではなく、任意の構造です。
細菌の細胞壁の化学組成:
- ペプチドグリカンムレイン;
- 多糖類;
- 脂質;
- タンパク質。
通常、この構造には外側と内側の 2 層があります。 植物と同じ働きをします。 ボディの一定の形状を維持および定義し、機械的保護を提供します。

教育紛争
私たちは細菌細胞の構造をある程度詳しく調べてきました。 バクテリアがどのようにして生き残れるかについて言及するだけが残っています 有利な条件、非常に長い間生存能力を失うことなく。
彼らは紛争と呼ばれる構造を形成することによってこれを行います。 それは生殖とは何の関係もなく、細菌を不利な条件から守るだけです。 紛争の形態はさまざまです。 通常の環境条件が回復すると、胞子が発生し、活性な細菌に成長します。
原核生物の細胞は、複雑で厳密に秩序だった構造を持ち、超微細構造組織とその基本的な特徴を備えています。 化学組成.
細菌細胞の構造構成要素は、基本的な構成要素と一時的な構成要素に分けられます (図 2)。 主な構造は、細胞壁、細胞質膜とその誘導体、細胞質とリボソームおよびさまざまな封入体、核様体です。 一時的 - 莢膜、粘膜、鞭毛、絨毛、内生胞子は、細菌の生活環の特定の段階でのみ形成され、一部の種ではそれらは完全に存在しません。
原核細胞では、細胞質膜の外側にある構造は表面(細胞壁、莢膜、鞭毛、絨毛)と呼ばれます。
「エンベロープ」という用語は、現在、細菌の細胞壁および莢膜、または細胞壁のみを指すのに使用されており、細胞質膜はエンベロープの一部ではなく、プロトプラストを指します。
細胞壁は細菌細胞の重要な構造要素であり、細胞質膜と莢膜の間に位置します。 非莢膜細菌では、それは外側の細胞膜です。 マイコプラズマと L 型細菌を除くすべての原核生物には必須です。 多くの機能を実行します。細菌を浸透圧ショックやその他の有害な要因から保護し、細菌の形状を決定し、代謝に参加します。 多くの種類の病原性細菌では、これは毒性があり、表面抗原を含み、表面にファージに対する特異的な受容体も持っています。 細菌の細胞壁には、外毒素やその他の細菌外タンパク質の輸送に関与する細孔が含まれています。 細胞壁の厚さは10~100nmで、細胞の乾物の5~50%を占めます。
細菌細胞壁の主成分はペプチドグリカン、またはムレイン(ラテン語で壁)であり、ネットワーク構造を持ち、細菌細胞の硬い(硬い)外枠を形成する支持ポリマーです。 ペプチドグリカンは、1,4-グリコシド結合によって接続された交互の N-アセチル-M-グルコサミンと N-アセチルムラミン酸残基、N-アセチルムラミン酸分子に結合した同一のテトラペプチド側鎖、および短いクロスペプチドからなる主鎖 (バックボーン) を持っています。鎖 多糖鎖をつなぐ橋。 ペプチドグリカンのサブユニットを接続する 2 種類の結合 (グリコシド結合とペプチド結合) により、このヘテロポリマーに分子ネットワーク構造が与えられます。 ペプチドグリカン層のコアはすべての細菌種で同じです。 テトラペプチドタンパク質鎖とペプチド (横) 鎖は、種によって異なります。
着色特性に基づいて、すべての細菌はグラム陽性菌とグラム陰性菌の 2 つのグループに分類されます。 1884 年、H. グラムは細菌を区別するために使用される染色法を提案しました。 この方法の本質は、グラム陽性菌がリンドウの紫とヨウ素の複合体をしっかりと固定し、エタノールによる漂白を受けないため、追加の色素フクシンを認識せず、紫色のままであることです。 グラム陰性菌では、この複合体はエタノールによって細胞から簡単に洗い流され、さらにフクシンを加えると赤くなります。 一部の細菌では、活発な増殖段階でのみグラム染色陽性が観察されます。 原核生物がグラム染色またはエタノールで脱色できるかどうかは、原核生物の細胞壁の特定の化学組成と超微細構造によって決まります。 グラム陽性菌のペプチドグリカンは細胞壁の主成分で 50 ~ 90% を占め、グラム陰性菌では 1 ~ 10% を占めます。 グラム陰性菌のペプチドグリカンの構造ミクロフィブリルはそれほど緻密に架橋されていないため、そのペプチドグリカン層の細孔はグラム陽性菌の分子骨格よりもはるかに広いです。 このようなペプチドグリカンの構造構成により、グラム陰性菌内のゲンチアナバイオレットとヨウ素のバイオレット複合体はより速く洗い流されます。
グラム陽性菌の細胞壁は細胞質膜にしっかりと隣接しており、大きく、その厚さは 20 ~ 100 nm の範囲にあります。 それはテイコ酸の存在を特徴とし、ペプチドグリカンと結合しており、三価アルコール - グリセロールまたは五原子アルコール - リビトールのポリマーであり、その残基はホスホジエステル結合によって結合されています。 テイコ酸はマグネシウムイオンと結合し、細胞内へのマグネシウムイオンの輸送に関与します。 グラム陽性原核生物の細胞壁には、多糖類、タンパク質、脂質も少量含まれています。
米。 2. 原核細胞の構造図:
1 - カプセル; 2 - 細胞壁。 3 - 細胞質膜。 4 - 核様体。 5 - 細胞質。 6 - 発色団。 7 - チラコイド。 8 - メソソーマ。 9 - リボソーム。 10 - 鞭毛。 11 - 基礎本体。 12 - 飲んだ。 13 - 硫黄の含有。 14 - 脂肪の滴。 15 - ポリリン酸顆粒。 16 - プラスミド
グラム陰性菌の細胞壁は多層になっており、その厚さは14~17nmです。 内層はペプチドグリカンであり、細胞を取り囲む薄い (2 nm) 連続ネットワークを形成します。 ペプチドグリカンにはメソジアミノピメリン酸のみが含まれ、リジンは含まれません。 細胞壁の外層、つまり外膜は、リン脂質、リポ多糖、リポタンパク質、タンパク質で構成されています。 外膜にはマトリックスタンパク質が含まれており、ペプチドグリカン層にしっかりと結合しています。 それらの機能の 1 つは、膜内に親水性の細孔を形成し、そこを通じて最大 600、場合によっては 900 の質量を持つ分子の拡散が起こることです。さらに、マトリックスタンパク質は、一部のファージの受容体としても機能します。 グラム陰性菌の細胞壁にあるリポ多糖類 (LPS) は、リピド A と多糖類から構成されています。 LPSは動物にとって有毒であり、エンドトキシンと呼ばれます。 テイコ酸はグラム陰性菌では見つかっていません。
グラム陰性菌の細胞壁の構造成分は細胞膜から境界を定められており、ペリプラズムまたはペリプラズム空間と呼ばれる空間によって分離されています。
プロトプラストとスフェロプラスト。 プロトプラストは細胞壁を完全に欠いた原核生物の形態であり、通常はグラム陽性細菌で形成されます。 スフェロプラストは、細胞壁が部分的に破壊された細菌です。 それらは外膜の要素を保持します。 それらはグラム陰性菌で観察されますが、グラム陽性菌ではそれほど頻繁ではありません。 これらは、適切な浸透圧の環境下で、リゾチームなどの溶解酵素によるペプチドグリカン層の破壊、または抗生物質ペニシリンなどによるペプチドグリカンの生合成の阻害の結果として形成されます。
プロトプラストおよびスフェロプラストは球状または半球状の形状をしており、元の細胞よりも 3 ~ 10 倍大きいです。 通常の条件下では、浸透圧溶解が起こり、死滅します。 浸透圧が上昇した条件下では、それらはしばらくの間生存し、成長し、さらには分裂することができます。 ペプチドグリカンを破壊する因子が除去されると、通常、プロトプラストは死滅しますが、L 型に変化する可能性があります。 スフェロプラストは容易に元の細菌に戻り、時には L 型に変化したり、死滅したりすることがあります。
L 型細菌。 これらは、細胞壁ペプチドグリカンを合成する能力を部分的または完全に失った細菌の表現型の改変、または突然変異体です。 したがって、L 型は細胞壁に欠陥のある細菌です。 それらは、1935年に英国のリスター研究所で分離され記載されたという事実に基づいてその名前が付けられました。それらは、抗生物質(ペニシリン、ポリミキシン、バシトラシン、ベンコマイシン、ストレプトマイシン)、アミノ酸などのL形質転換剤の影響下で形成されます。 (グリシン、メチオニン、ロイシンなど)、酵素リゾチーム、紫外線、X 線。 プロトプラストやスフェロプラストとは異なり、L 型は比較的高い生存率と顕著な繁殖能力を持っています。 形態学的および文化的特性の点で、それらは元の細菌とは大きく異なります。これは、細胞壁の喪失と代謝活性の変化によるものです。
L 型細菌には多型があります。 0.2〜1ミクロンの基本天体(最小生殖要素)、球体 - 1〜5、大きな天体 - 5〜50、糸 - 最大4ミクロン以上があります。 L 型細胞は、細胞質内膜とミエリン様構造のよく発達したシステムを持っています。 細胞壁の欠陥により、それらは浸透圧的に不安定であり、浸透圧の高い特別な培地でのみ培養できます。 それらはバクテリアフィルターを通過します。
細菌には安定した L 型と不安定な L 型が存在します。 前者は硬い細胞壁をまったく持たないため、プロトプラストに似ています。 元の細菌の形態に戻ることはほとんどありません。 後者は細胞壁の要素を持っている可能性があり、スフェロプラストに似ています。 形成を引き起こした因子が存在しない場合、それらは元の細胞に戻ります。
L 型の形成プロセスは、L 変換または L 誘導と呼ばれます。 病原性細菌(ブルセラ症、結核、リステリア菌などの原因物質)を含むほぼすべての種類の細菌は、L 型変換を受ける能力を持っています。
L 型は、慢性再発性感染症の発症、病原体の保有、体内での長期残留において非常に重要視されています。 L 型細菌の素体の経胎盤侵入性は証明されています。
L型細菌によって引き起こされる感染過程は、非定型性、経過期間、疾患の重症度を特徴とし、化学療法による治療が困難です。
莢膜は細菌の細胞壁の上にある粘膜層です。 カプセル物質は明確に区別されています。 環境。 層の厚さと細菌細胞との結合の強さに応じて、厚さ0.2ミクロンを超えるマクロカプセルは光学顕微鏡ではっきりと見え、マイクロカプセルは厚さ0.2ミクロン未満でのみ検出可能です。電子顕微鏡による検出、または化学的および免疫学的方法による検出が区別されます。 マクロカプセル (真のカプセル) は、B. anlhracis、C1 によって形成されます。 ウェルシュ菌、マイクロカプセル - Escherichia coJi。 カプセルは細菌細胞の必須の構造ではありません。カプセルが失われても細菌は死に至りません。 細菌の莢膜を持たない突然変異体、例えば炭疽菌ワクチン株STI-1が知られている。
カプセルの物質は親水性の高いミセルで構成されており、その化学組成は非常に多様です。 ほとんどの原核生物のカプセルの主成分はホモ多糖またはヘツロ多糖 (腸内細菌など) です。 一部の種類の桿菌では、カプセルはポリペプチドから構築されます。 したがって、炭疽菌の莢膜の組成には、D-グルタミン酸ポリペプチド(右旋性異性体)が含まれる。 哺乳類の結核菌のマイクロカプセルの組成には、トレハロースとミコール酸のエステル(コードファクター)に代表される糖ペプチドが含まれています。
カプセル合成は複雑なプロセスであり、原核生物ごとに独自の特徴があります。 カプセル生体高分子は細胞質膜の外表面で合成され、細胞壁の特定の領域の表面に放出されると考えられています。
粘液を合成する細菌がいますが、粘液は多糖類の構造のない層の形で細胞壁の表面に堆積します。 細胞を取り囲む粘液物質は、多くの場合、細胞の直径よりも厚いです。 腐生細菌 Leuconostoca では、多くの個体に対して 1 つの莢膜の形成が観察されます。 共通のカプセル内に封入されたこのような細菌の蓄積は、ズーグレアと呼ばれます。
カプセルは、重要な生物学的役割を果たす多機能細胞小器官です。 これは、細菌の毒性、抗原特異性、および免疫原性を決定する莢膜抗原の局在部位です。 病原菌における莢膜の喪失は、例えば炭疽菌の非莢膜株において、その毒性を急激に減少させる。 カプセルは細菌の生存を確保し、機械的損傷、乾燥、ファージ、有毒物質による感染、および病原性形態から細菌を保護します。つまり、カプセル化された細胞は貪食されにくいです。 病原性細菌を含む一部の種類の細菌では、基質への細胞の付着を促進します。
獣医学微生物学では、炭疽菌を検査する際に莢膜の検出が病原体の形態学的徴候として使用されます。
カプセルの着色には、ロマノフスキー、ギムザ、ジン、ブリ、オルト、ミヒンなどの特別な方法が使用されます。
マイクロカプセルと粘膜層は血清学的反応 (RA) によって測定され、カプセルの抗原成分は免疫蛍光法 (RIF) と RDD を使用して同定されます。
鞭毛は細菌の運動を行う細胞小器官であり、タンパク質の性質を持った細くて長い糸状の構造で表されます。 その長さは細菌細胞の数倍を超え、10〜20ミクロンであり、一部のスピリラでは80〜90ミクロンに達します。 鞭毛フィラメント (フィブリル) は、直径 12 ~ 20 nm の完全ならせん状の円柱です。 ビブリオスとプロテウスでは、フィラメントは厚さ 35 nm の鞘で囲まれています。
鞭毛は、らせん状フィラメント、フック、基部の 3 つの部分で構成されています。 フックは湾曲したタンパク質のシリンダーであり、基底体と鞭毛の硬いフィラメントの間の柔軟なリンクとして機能します。 基底体は中心の棒(軸)とリングからなる複雑な構造です。

米。 3.鞭毛:
a - モノリッチ。 b - アンフィトリク。 c - ロフォトリクス。 d - 周膜質
鞭毛は細菌細胞の重要な構造ではありません。細菌には段階の変化があり、細胞発生のある段階では存在し、別の段階では存在しません。 したがって、古い培養物における破傷風の原因物質では、鞭毛を持たない細胞が優勢です。
異なる種の細菌における鞭毛の数(1 個から 50 本以上)とその局在場所は同じではありませんが、1 つの種では安定しています。 これに応じて、以下の鞭毛細菌のグループが区別されます。 両親媒性 - 極性を持って配置された 2 本の鞭毛を持つ細菌、または両端に鞭毛の束を持つ細菌。 ロフォトリクス - 細胞の一端に鞭毛の束を持つ細菌。 ペリトリクは、細胞の側面または表面全体に多数の鞭毛を持つ細菌です(図3)。 鞭毛を持たない細菌は無毛細菌と呼ばれます。
運動器官である鞭毛は、浮遊する棒状の複雑な形の細菌の典型であり、球菌の孤立した場合にのみ見られます。 これらは、液体媒体中では効率的な動きを提供し、固体基板の表面ではより遅い動きを提供します。 単三毛虫と偏毛虫の移動速度は 50 μm/s に達しますが、両毛虫と周毛虫はよりゆっくりと移動し、通常は細胞のサイズに等しい距離を 1 秒で移動します。
細菌はランダムに移動しますが、外部刺激によって決定される、指示された形式の移動、つまりタクシーが可能です。 さまざまな環境要因に反応して、細菌は短時間で最適な生息域に集中します。 タクシーにはプラスの場合もマイナスの場合もあります。 走化性、気流性、光走性、磁走性を区別するのが通例です。 走化性は環境内の化学物質の濃度の違いによって引き起こされ、エアロタキシスは酸素によって、走光性は光の強度によって引き起こされ、磁気走性は微生物が磁場内を移動する能力によって決定されます。
細菌の運動性鞭毛形態の同定は、感染症の臨床検査における細菌の同定にとって重要です。
線毛 (線毛、絨毛) は、厚さ 3 ~ 25 nm、長さ 12 μm までの真っ直ぐで薄い中空のタンパク質のシリンダーで、細菌細胞の表面から伸びています。 それらは特定のタンパク質であるピリンによって形成され、細胞質膜に由来し、細菌の運動性および不動性の形態で見られ、電子顕微鏡でのみ見ることができます(図4)。 細胞の表面には、1〜2、50〜400またはそれ以上から数千の線毛が存在する可能性があります。

米。 4. 飲酒
線毛には 2 つのクラスがあります: 性線毛 (sexpili) と一般線毛 (より多くの場合線毛と呼ばれます)。 同じ細菌でも異なる性質の線毛を持つことがあります。 性線毛は、接合の過程で細菌の表面に現れ、遺伝物質(DNA)がドナーからレシピエントに伝達される細胞小器官の機能を果たします。
一般的なタイプの線毛は、毛束周囲(大腸菌)または極(シュードモナス)に位置します。 1 つの細菌には何百もの細菌が含まれている場合があります。 それらは、細菌の凝集体への付着、細胞を含むさまざまな基質への微生物の付着(付着機能)、代謝産物の輸送に関与し、液体媒体の表面での膜の形成にも寄与します。 赤血球の凝集を引き起こします。
細胞質膜とその誘導体。 細胞質膜 (原形質膜) は、細胞壁から細胞質を分離する細菌細胞の半透過性リポタンパク質構造です。 これは細胞の必須の多機能成分であり、乾燥質量の 8 ~ 15% を占めます。 細胞質膜の破壊は細菌細胞の死をもたらします。 電子顕微鏡での極薄切片では、その 3 層構造が明らかになります。厚さ 2 ~ 3 nm の 2 つの限定的な親浸透性層と、厚さ 4 ~ 5 nm の 1 つの疎浸透性中心層です。
化学的には、細胞質膜は 50 ~ 75% のタンパク質と 15 ~ 50% の脂質からなるタンパク質 - 脂質複合体です。 膜脂質の主要部分 (70 ~ 90%) はリン脂質で表されます。 それは 2 つの単分子タンパク質層から構築されており、その間に規則正しく配向した 2 列の脂質分子からなる脂質層があります。
細胞質膜は細胞に対する浸透圧障壁として機能し、細胞内への栄養素の流れと外部への代謝産物の放出を制御します。細胞質膜には、有機分子および無機分子の能動的選択的移動を実行する基質特異的透過酵素が含まれています。
細胞質膜酵素は、膜脂質、細胞壁成分、カプセルおよび外酵素の合成の最終段階を触媒します。 エネルギー合成を担う酸化的リン酸化酵素と電子伝達酵素は膜上に局在しています。
細胞増殖中に、細胞質膜は細胞質内膜構造を形成する多数の陥入を形成します。 局所的な膜の陥入はメソソームと呼ばれます。 これらの構造は、グラム陽性菌ではよく発現されますが、グラム陰性菌では発現が悪く、リケッチアやマイコプラズマではあまり発現されません。
メソソームと細菌の染色体との間の接続が確立されており、そのような構造はヌクレオイドソームと呼ばれます。 核様体と統合されたメソソームは核分裂と細胞質分裂に関与します 微生物細胞、DNA複製の終了後のゲノムの分布とその後の娘染色体の分岐を確実にします。 メソソームは、細胞質膜と同様、細菌の呼吸活動の中心であるため、ミトコンドリアの類似体と呼ばれることもあります。 しかし、メソソームの重要性はまだ完全には解明されていません。 それらは膜の作用面を増加させ、おそらく構造的機能のみを実行し、細菌細胞を比較的別々の区画に分割し、酵素プロセスの発生にとってより好ましい条件を作り出します。 病原性細菌では、それらは外毒素のタンパク質分子の輸送を確実にします。
細胞質は細菌細胞の内容物であり、細胞質膜によって区切られています。 それは、可溶性RNA成分、基質物質、酵素、代謝産物、および構造要素であるリボソーム、細胞質内膜、封入体および核様体を含む均一な画分であるサイトゾルで構成されています。
リボソームは、タンパク質の生合成を実行する細胞小器官です。 それらはタンパク質と RNA で構成されており、水素結合と疎水結合によって複合体を形成しています。 細菌のリボソームは直径 15 ~ 20 nm の顆粒で、沈降定数は 70S で、2 つのリボ核タンパク質サブユニット、30S と 50S から形成されます。 1 つの細菌細胞には 5,000 ~ 50,000 個のリボソームが含まれており、mRNA を介して結合して、高いタンパク質合成活性を持つ 50 ~ 55 個のリボソームからなるポリソーム凝集体が形成されます。
細菌の細胞質にはさまざまなタイプの封入体が検出されます。 それらは、タンパク質膜の有無にかかわらず、固体、液体、または気体であり、永久的に存在するわけではありません。 それらの重要な部分は、予備栄養素と細胞代謝産物です。 埋蔵栄養素には、多糖類、脂質、ポリリン酸塩、硫黄堆積物などが含まれます。多糖類の性質を持つ含有物の中で、グリコーゲンとデンプン様物質顆粒膜が最も多く見られ、これらは炭素およびエネルギー物質の供給源として機能します。 脂質は顆粒および脂肪滴の形で細胞内に蓄積します。これらには、光を鋭く屈折させ、光学顕微鏡ではっきりと見えるポリ-β-ヒドロキシ酪酸の膜に囲まれた顆粒が含まれます。 炭疽菌や好気性芽胞形成腐生菌も検出されます。 マイコバクテリアは予備物質としてワックスを蓄積します。 一部の麻疹非細菌、スピリラなどの細胞には、ポリリン酸によって形成されるボルチン顆粒が含まれています。 異色症が特徴で、トルイジンブルーとメチレンブルーで紫赤色に染まります。 ボルチン顆粒はリン酸貯蔵庫の役割を果たします。
膜に囲まれた封入体には、気液胞またはエアロソームも含まれます。これらは細胞の比重を軽減し、水生原核生物に見られます。
核様体は原核生物の核です。 これは、長さ 1.1 ~ 1.6 nm の環状に閉じた 1 本の二本鎖 DNA 鎖で構成されており、単一の細菌染色体またはジェノフォアと考えられています。
原核生物の核様体は膜によって細胞の他の部分と区切られておらず、核膜がありません。
核様体構造には、RNA ポリメラーゼ、塩基性タンパク質、欠損ヒストンが含まれます。 染色体は細胞質膜に固定されており、グラム陽性菌ではメソソームに固定されています。 細菌の染色体は多保存的な方法で複製します。つまり、親 DNA 二重らせんがほどかれ、新しい相補鎖が各ポリヌクレオチド鎖の鋳型上に組み立てられます。 核様体には有糸分裂装置がなく、娘核の分離は細胞質膜の成長によって確実に行われます。
細菌のコアは分化した構造です。 細胞発生の段階に応じて、核様体は個別(不連続)であり、個々の断片で構成される場合があります。 これは、細菌細胞の分裂が DNA 分子の複製サイクルの完了と娘染色体の形成後に起こるという事実によるものです。
核様体には、細菌細胞の遺伝情報の大部分が含まれています。
核様体に加えて、多くの細菌の細胞には染色体外遺伝要素、つまり自律複製可能な小さな環状 DNA 分子であるプラスミドが見られます。