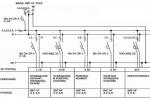
Структура бактериальной клетки
Структура бактерий хорошо изучена с помощью электронной микроскопии целых клеток и их ультратонких срезов. Бактериальная клетка состоит из клеточной стенки, цитоплазматической мембраны, цитоплазмы с включениями и ядра, называемого нуклеоидом. Имеются дополнительные структуры: капсула, микрокапсула, слизь, жгутики, пили (рис.1); некоторые бактерии в неблагоприятных условиях способны образовывать споры.
Клеточная стенка - прочная, упругая структура, придающая бактерии определенную форму и вместе с подлежащей цитоплазматической мембраной «сдерживающая» высокое осмотическое давление в бактериальной клетке. Она участвует в процессе деления клетки и транспорте метаболитов. Наиболее толстая клеточная стенка у грамположительных бактерий (рис.1). Так, если толщина клеточной стенки грамотрицательных бактерий около 15-20 нм, то у грамположительных она может достигать 50 нм и более. В клеточной стенке грамположительных бактерий содержится небольшое количество полисахаридов, липидов, белков.
Основным компонентом клеточной стенки этих бактерий является многослойный пептидогликан (муреин, мукопептид), составляющий 40-90 % массы клеточной стенки.
Волютин Мезосома Нуклеоид
Рис. 1. Строение бактериальной клетки.
С пептидогликаном клеточной стенки грамположительных бактерий ковалентно связаны тейхоевые кислоты (от греч. teichos - стенка), молекулы которых представляют собой цепи из 8-50 остатков глицерола и рибитола, соединенных фосфатными мостиками. Форму и прочность бактериям придает жесткая волокнистая структура многослойного с поперечными пептидными сшивками пептидогликана. Пептидогликан представлен параллельно расположенными молекулами гликана, состоящего из повторяющихся остатков N -ацетилглюкозамина и N -ацетилмурамовой кислоты, соединенных гликозидной связью типа Р (1 -> 4).
Лизоцим, являясь ацетилмурамидазой, разрывает эти связи. Гликановые молекулы связаны поперечной пептидной связью. Отсюда и название этого полимера - пептидогликан. Основу пептидной связи пептидогликана грамотрицательных бактерий составляют тетрапептиды, состоящие из чередующихся L- и Д -аминокислот.
У E. coli пептидные цепи соединены друг с другом через D- аланин одной цепи и мезодиаминопимелиновую кислоту другой.
Состав и строение пептидной части пептидогликана у грамотрицательных бактерий стабильны в отличие от пептидогликана грамположительных бактерий, аминокислоты которого могут отличаться по составу и последовательности. Тетрапептиды здесь соединены друг с другом полипептидными цепочками из 5 остатков глицина. У грамположительных бактерий вместо мезодиаминопимелиновой кислоты часто содержится лизин. Фосфолипид

Рис. 2. Строение поверхностных структур грамположительных (грам+) и грамотрицательных (грам") бактерий.
Элементы гликана (ацетилглюкозамин и ацетилмурамовая кислота) и аминокислоты тетрапептида (мезодиаминопимелиновая и Л-глутаминовая кислоты, Д-аланин) являются отличительной особенностью бактерий, поскольку они и Д-изомеры аминокислот отсутствуют у животных и человека.
Способность грамположительных бактерий при окраске по Граму удерживать генциановый фиолетовый в комплексе с йодом (сине-фиолетовая окраска бактерий) связана со свойством многослойного пептидогликана взаимодействовать с красителем. Кроме этого, последующая обработка мазка бактерий спиртом вызывает суживание пор в пептидогликане и тем самым задержку красителя в клеточной стенке. Грамотрицательные бактерии после воздействия спиртом утрачивают краситель, обесцвечиваются и при обработке фуксином окрашиваются в красный цвет. Это обусловлено меньшим количеством пептидогликана (5-10 % массы клеточной стенки).
В состав клеточной стенки грамотрицательных бактерий входит наружная мембрана, связанная посредством липопротеина с подлежащим слоем пептидогликана (рис.2). Наружная мембрана представляет собой волнообразную трехслойную структуру, сходную с внутренней мембраной, которую называют ци-топлазматической. Основным компонентом этих мембран служит бимолекулярный (двойной) слой липидов.
Наружная мембрана является асимметричной мозаичной структурой, представленной липополисахаридами, фосфолипидами и белками . С ее внешней стороны расположен липополисахарид (ЛПС), состоящий из трех компонентов: липида А, стержневой части, или ядра (лат. core - ядро), и 0-специфической цепи полисахарида, образованной повторяющимися олигосахаридными последовательностями.
Липополисахарид «заякорен» в наружной мембране липидом А, обусловливающим токсичность ЛПС, отождествляемому поэтому с эндотоксином . Разрушение бактерий антибиотиками приводит к освобождению большого количества эндотоксина,чтоможет привести к эндотоксическому шоку больного.
От липида А отходит ядро, или стержневая часть ЛПС. Наиболее постоянной частью ядра ЛПС является кетодезоксиоктоновая кислота (3-деокси-г)-манно-2-октулосоновая кислота). 0 -специфическая цепь, отходящая от стержневой части молекулы ЛПС, обусловливает серогруппу, серовар (разновидность бактерий, выявляемая с помощью иммунной сыворотки) определенного штамма бактерий. Таким образом, с понятием ЛПС - связаны представления об 0-антигене, покоторому можно дифференцировать бактерии. Генетические изменения могут привести к изменениям в биосинтезе компонентовЛПС бактерий и к появлению в результате этого L -форм.
Белки матрикса наружной мембраны пронизывают ее таким образом, что молекулы белка, называемые поринами, окаймляют гидрофильные поры, через которые проходят вода и мелкие молекулы с относительной массой до 700. Между наружной и цитоплазматической мембранами находится периплазматическое пространство, или периплазма, содержащая ферменты. При нарушении синтеза клеточной стенки бактерий под влиянием лизоцима, пенициллина, защитных факторов организма и других соединений образуются клетки с измененной (часто шаровидной) формой: протопласты - бактерии, полностью лишенные клеточной стенки; сферопласты - бактерии с частично сохранившейся клеточной стенкой. После удаления ингибитора клеточной стенки такие измененные бактерии могут реверсировать, т.е. приобретать полноценную клеточную стенку и восстанавливать исходную форму.
Бактерии сферо- или протопластного типа, утратившие способность к синтезу пептидогликана под влиянием антибиотиков или других факторов и способные размножаться, называются L-формами (от названия института им. Листера). L -формы могут возникать и в результате мутаций. Они представляют собой осмотически чувствительные, шаровидные, колбовидные клетки различной величины, в том числе и проходящие через бактериальные фильтры. Некоторые L -формы (нестабильные) при удалении фактора, приведшего к изменениям бактерий, могут реверсировать, «возвращаясь» в исходную бактериальную клетку. L -формы могут образовывать многие возбудители инфекционных болезней.
Цитоплазматическая мембрана при электронной микроскопии ультратонких срезов представляет собой трехслойную мембрану, окружающую наружную часть цитоплазмы бактерий. По структуре она похожа на плазмалемму клеток животных и состоит из двойного слоя липидов, главным образом фосфолипидов с внедренными поверхностными, а также интегральными белками, как бы пронизывающими насквозь структуру мембраны. Некоторые из них являются пермеазами, участвующими в транспорте веществ. Цитоплазматическая мембрана является динамической структурой с подвижными компонентами, поэтому ее представляют как мобильную текучую структуру. Она участвует в регуляции осмотического давления, транспорте веществ и энергетическом метаболизме клетки (за счет ферментов цепи переноса электронов, аденозинтрифосфатазы и др.). При избыточном росте (по сравнению с ростом клеточной стенки) Цитоплазматическая мембрана образует инвагинаты - впячивания в виде сложно закрученных мембранных структур, называемые мезосомами. Менее сложно закрученные структуры называются внутрицитоплазматическими мембранами. Роль мезосом и внутрицитоплазматических мембран до конца не выяснена. Предполагают даже, что они являются артефактом, возникающим после приготовления (фиксации) препарата для электронной микроскопии. Тем не менее считают, что производные цитоплазматической мембраны участвуют в делении клетки, обеспечивая энергией синтез клеточной стенки, принимают участие в секреции веществ, спорообразовании, т.е. в процессах с высокой затратой энергии.
Цитоплазма занимает основной объем бактериальной клетки и состоит из растворимых белков, рибонуклеиновых кислот, включений и многочисленных мелких гранул - рибосом, ответственных за синтез (трансляцию) белков. Рибосомы бактерий имеют размер около 20 нм и коэффициент седиментации 70S, 3 отличие от 80^-рибосом, характерных для эукариотических клеток. Поэтому некоторые антибиотики, связываясь с рибосо-мами бактерий, подавляют синтез бактериального белка, не влияя на синтез белка эукариотических клеток. Рибосомы бактерий могут диссоциировать на две субъединицы - 50S и 30S . В цитоплазме имеются различные включения в виде гранул гликогена, полисахаридов, поли-р-масляной кислоты и полифосфатов (волютин). Они накапливаются при избытке питательных веществ в окружающей среде и выполняют роль запасных веществ для питания и энергетических потребностей. Волютин обладает сродством к основным красителям, обладает метахро-мазией и легко выявляется с помощью специальных методов окраски. Характерное расположение зерен волютина выявляется у дифтерийной палочки в виде интенсивно прокрашивающихся полюсов клетки.
Нуклеоид - эквивалент ядра у бактерий. Он расположен в центральной зоне бактерий в виде двунитчатой ДНК, замкнутой в кольцо и плотно уложенной наподобие клубка. В отличие от эукариот ядро бактерий не имеет ядерной оболочки, ядрышка и основных белков (гистонов). Обычно в бактериальной клетке содержится одна хромосома, представленная замкнутой в кольцо молекулой ДНК. При нарушении деления в ней может находиться 4 и более хромосом. Нуклеоид выявляется в световом микроскопе после окраски специфическими для ДНК методами: по Фельгену или по Романовскому-Гимзе. На электронограммах ультратонких срезов бактерий нуклеоид имеет вид светлых зон с фибриллярными, нитевидными структурами ДНК, связанной определенными участками с цитоплазматической мембраной или мезосомой, участвующими в репликации хромосомы.
Кроме нуклеоида, представленного одной хромосомой, в бактериальной клетке имеются внехромосомные факторы наследственности - плазмиды , представляющие собой ковалентно замкнутые кольца ДНК.
Капсула - слизистая структура толщиной более 0,2 мкм, прочно связанная с клеточной стенкой бактерий и имеющая четко очерченные внешние границы. Капсула различима в мазках-отпечатках из патологического материала. В чистых культурах бактерий капсула образуется реже. Она выявляется при специальных методах окраски по Бурри-Гинсу, создающих негативное контрастирование веществ капсулы.
Обычно капсула состоит из полисахаридов (экзополисахаридов), иногда из полипептидов, например у сибиреязвенной бациллы. Капсула гидрофильна, она препятствует фагоцитозу бактерий.
Многие бактерии образуют микрокапсулу - слизистое образование толщиной менее 0,2 мкм, выявляемое лишь при электронной микроскопии. От капсулы следует отличать слизь - мукоидные экзополисахариды, не имеющие четких внешних границ. Мукоидные экзополисахариды характерны для мукоидных штаммов синегнойной палочки, часто встречающихся в мокроте больных с кистозным фиброзом. Бактериальные экзополисахариды участвуют в адгезии (прилипании к субстратам), их еще называют гликокаликсом. Кроме синтеза экзополисахаридов бактериями, существует и другой механизм их образования: путем действия внеклеточных ферментов бактерий на дисахариды. В результате этого образуются декстраны и леваны. Капсула и слизь предохраняют бактерии от повреждений, высыхания, так как, являясь гидрофильными, хорошо связывают воду, препятствуют действию защитных факторов макроорганизма и бактериофагов.
Жгутики бактерий определяют подвижность бактериальной клетки. Жгутики представляют собой тонкие нити, берущие начало от цитоплазматической мембраны, имеют большую длину, чем сама клетка (рис.3). Толщина жгутиков 12-20 нм, длина 3-12 мкм. Число жгутиков у бактерий различных видов варьирует от одного (монотрих) у холерного вибриона до десятка и сотен жгутиков, отходящих по периметру бактерии (пери-трих) у кишечной палочки, протея и др. Лофотрихи имеют пучок жгутиков на одном из концов клетки. Амфитрихи имеют по одному жгутику или пучку жгутиков на противоположных концах клетки. Жгутики прикреплены к цитоплазматической мембране и клеточной стенке специальными дисками. Жгутики состоят из белка - флагеллина (от naT.flagellum - жгутик), обладающего антигенной специфичностью. Субъединицы флагеллина закруче ны в виде спирали. Жгутики выявляют с помощью электронной микроскопии препаратов, напыленных тяжелыми металлами, или в световом микроскопе после обработки специальными методами, основанными на протравливании и адсорбции различных веществ, приводящих к увеличению толщины жгутиков (например, после серебрения).

Рис. 3. Кишечная палочка. Электронограмма (препарат В.С.Тюрина). 1 - жгутики, 2 - ворсинки, 3 - F-пили.
Ворсинки, или пили (фимбрии), - нитевидные образования (рис.3), более тонкие и короткие (3-10 нм х 0,3-10 мкм), чем жгутики. Пили отходят от поверхности клетки и состоят из белка пилина. Они обладают антигенной активностью. Среди пилей выделяются: пили, ответственные за адгезию, т.е. за прикрепление бактерий к поражаемой клетке (пили 1-го типа, или общего типа - common pili), пили, ответственные за питание, водно-солевой обмен; половые (F-пили), или конъюга-ционные пили (пили 2-го типа). Пили общего типа многочисленны - несколько сотен на клетку. Половые пили образуются так называемыми «мужскими» клетками-донорами, содержащими трансмиссивные плазмиды (F, R, Col). Их обычно бывает 1-3 на клетку. Отличительной особенностью половых пилей является взаимодействие с особыми «мужскими» сферически ми бактериофагами, которые интенсивно адсорбируются на половых пилях.
Споры - своеобразная форма покоящихся фирмикутных бактерий, т.е. бактерий с грамположительным типом строения клеточной стенки.
Споры образуются при неблагоприятных условиях существования бактерий (высушивание, дефицит питательных веществ и др.). При этом внутри одной бактерии образуется одна спора. Образование спор способствует сохранению вида и не является способом размножения, как у грибов.
Спорообразующие аэробные бактерии, у которых размер споры не превышает диаметр клетки, иногда называются бациллами. Спорообразующие анаэробные бактерии, у которых размер споры превышает диаметр клетки, и поэтому они принимают форму веретена, называются клостридиями (лат. clostridium - веретено).
Процесс спорообразования (споруляция) проходит ряд стадий, в течение которых часть цитоплазмы и хромосома отделяются, окружаясь цито плазматической мембраной; образуется проспора, затем формируется многослойная плохо проницаемая оболочка. Спорообразование сопровождается интенсивным потреблением проспорой, а затем формирующейся оболочкой споры дипиколиновой кислоты и ионов кальция. После формирования всех структур спора приобретает термоустойчивость, которую связывают с наличием дипиколината кальция. Спорообразование, форма и расположение спор в клетке (вегетативной) являются видовым свойством бактерий, что позволяет отличать их друг от друга. Форма спор может быть овальной, шаровидной, расположение в клетке - терминальное, т.е. на конце палочки (возбудитель столбняка), субтерминальное - ближе к концу палочки (возбудители ботулизма, газовой гангрены) и центральное (сибиреязвенная бацилла).
Бактерии — самая древняя группа организмов из ныне существующих на Земле. Первые бактерии появились, вероятно, более 3,5 млрд лет назад и на протяжении почти миллиарда лет были единственными живыми существами на нашей планете. Поскольку это были первые представители живой природы, их тело имело примитивное строение.
Со временем их строение усложнилось, но и поныне бактерии считаются наиболее примитивными одноклеточными организмами. Интересно, что некоторые бактерии и сейчас ещё сохранили примитивные черты своих древних предков. Это наблюдается у бактерий, обитающих в горячих серных источниках и бескислородных илах на дне водоёмов.
Большинство бактерий бесцветно. Только немногие окрашены в пурпурный или в зелёный цвет. Но колонии многих бактерий имеют яркую окраску, которая обусловливается выделением окрашенного вещества в окружающую среду или пигментированием клеток.
Первооткрывателем мира бактерий был Антоний Левенгук — голландский естествоиспытатель 17 века, впервые создавший совершенную лупу-микроскоп, увеличивающую предметы в 160-270 раз.
Бактерии относят к прокариотам и выделяют в отдельное царство — Бактерии.
Форма тела
Бактерии — многочисленные и разнообразные организмы. Они различаются по форме.
| Название бактерии | Форма бактерии | Изображение бактерии |
| Кокки | Шарообразная | |
| Бацилла |  | Палочковидная |
| Вибрион | Изогнутая в виде запятой | |
| Спирилла |  | Спиралевидная |
| Стрептококки |  | Цепочка из кокков |
| Стафилококки |  | Грозди кокков |
| Диплококки | Две круглые бактерии, заключённые в одной слизистой капсуле |
Способы передвижения
Среди бактерий есть подвижные и неподвижные формы. Подвижные передвигаются за счёт волнообразных сокращений или при помощи жгутиков (скрученные винтообразные нити), которые состоят из особого белка флагеллина. Жгутиков может быть один или несколько. Располагаются они у одних бактерий на одном конце клетки, у других — на двух или по всей поверхности.

Но движение присуще и многим иным бактериям, у которых жгутики отсутствуют. Так, бактерии, покрытые снаружи слизью, способны к скользящему движению.
У некоторых лишённых жгутиков водных и почвенных бактерий в цитоплазме имеются газовые вакуоли. В клетке может быть 40-60 вакуолей. Каждая из них заполнена газом (предположительно — азотом). Регулируя количество газа в вакуолях, водные бактерии могут погружаться в толщу воды или подниматься на её поверхность, а почвенные бактерии — передвигаться в капиллярах почвы.
Место обитания
В силу простоты организации и неприхотливости бактерии широко распространены в природе. Бактерии обнаружены везде: в капле даже самой чистой родниковой воды, в крупинках почвы, в воздухе, на скалах, в полярных снегах, песках пустынь, на дне океана, в добытой с огромной глубины нефти и даже в воде горячих источников с температурой около 80ºС. Обитают они на растениях, плодах, у различных животных и у человека в кишечнике, ротовой полости, на конечностях, на поверхности тела.
Бактерии — самые мелкие и самые многочисленные живые существа. Благодаря малым размерам они легко проникают в любые трещины, щели, поры. Очень выносливы и приспособлены к различным условиям существования. Переносят высушивание, сильные холода, нагревание до 90ºС, не теряя при этом жизнеспособность.
Практически нет места на Земле, где не встречались бы бактерии, но в разных количествах. Условия жизни бактерий разнообразны. Одним из них необходим кислород воздуха, другие в нём не нуждаются и способны жить в бескислородной среде.
В воздухе: бактерии поднимаются в верхние слои атмосферы до 30 км. и больше.
Особенно много их в почве. В 1 г. почвы могут содержаться сотни миллионов бактерий.
В воде: в поверхностных слоях воды открытых водоёмов. Полезные водные бактерии минерализуют органические остатки.
В живых организмах: болезнетворные бактерии попадают в организм из внешней среды, но лишь в благоприятных условиях вызываю заболевания. Симбиотические живут в органах пищеварения, помогая расщеплять и усваивать пищу, синтезируют витамины.
Внешнее строение
Клетка бактерии одета особой плотной оболочкой — клеточной стенкой, которая выполняет защитную и опорную функции, а также придаёт бактерии постоянную, характерную для неё форму. Клеточная стенка бактерии напоминает оболочку растительной клетки. Она проницаема: через неё питательные вещества свободно проходят в клетку, а продукты обмена веществ выходят в окружающую среду. Часто поверх клеточной стенки у бактерий вырабатывается дополнительный защитный слой слизи — капсула. Толщина капсулы может во много раз превышать диаметр самой клетки, но может быть и очень небольшой. Капсула — не обязательная часть клетки, она образуется в зависимости от условий, в которые попадают бактерии. Она предохраняет бактерию от высыхания.
На поверхности некоторых бактерий имеются длинные жгутики (один, два или много) или короткие тонкие ворсинки. Длина жгутиков может во много раз превышать разметы тела бактерии. С помощью жгутиков и ворсинок бактерии передвигаются.

Внутреннее строение
Внутри клетки бактерии находится густая неподвижная цитоплазма. Она имеет слоистое строение, вакуолей нет, поэтому различные белки (ферменты) и запасные питательные вещества размещаются в самом веществе цитоплазмы. Клетки бактерий не имеют ядра. В центральной части их клетки сконцентрировано вещество, несущее наследственную информации. Бактерии, — нуклеиновая кислота — ДНК. Но это вещество не оформлено в ядро.
Внутренняя организация бактериальной клетки сложна и имеет свои специфические особенности. Цитоплазма отделяется от клеточной стенки цитоплазматической мембраной. В цитоплазме различают основное вещество, или матрикс, рибосомы и небольшое количество мембранных структур, выполняющих самые различные функции (аналоги митохондрий, эндоплазматической сети, аппарата Гольджи). В цитоплазме клеток бактерий часто содержатся гранулы различной формы и размеров. Гранулы могут состоять из соединений, которые служат источником энергии и углерода. В бактериальной клетке встречаются и капельки жира.
В центральной части клетки локализовано ядерное вещество — ДНК, не отграниченная от цитоплазмы мембраной. Это аналог ядра — нуклеоид. Нуклеоид не обладает мембраной, ядрышком и набором хромосом.
Способы питания
У бактерий наблюдаются разные способы питания. Среди них есть автотрофы и гетеротрофы. Автотрофы — организмы, способные самостоятельно образовывать органические вещества для своего питания.
Растения нуждаются в азоте, но сами усваивают азот воздуха не могут. Некоторые бактерии соединяют содержащиеся в воздухе молекулы азота с другими молекулами, в результате чего получаются вещества, доступные для растений.
Эти бактерии поселяются в клетках молодых корней, что приводит к образованию на корнях утолщений, называемых клубеньками. Такие клубеньки образуются на корнях растений семейства бобовых и некоторых других растений.
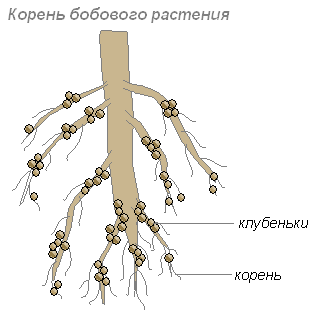
Корни дают бактериям углеводы, а бактерии корням — такие содержащие азот вещества, которые могут быть усвоены растением. Их сожительство взаимовыгодно.
Корни растений выделяют много органических веществ (сахара, аминокислоты и другие), которыми питаются бактерии. Поэтому в слое почвы, окружающем корни, поселяется особенно много бактерий. Эти бактерии превращают отмершие остатки растений в доступные для растения вещества. Этот слой почвы называют ризосферой.
Существует несколько гипотез о проникновении клубеньковых бактерий в ткани корня:
- через повреждения эпидермальной и коровой ткани;
- через корневые волоски;
- только через молодую клеточную оболочку;
- благодаря бактериям-спутникам, продуцирующим пектинолитические ферменты;
- благодаря стимуляции синтеза В-индолилуксусной кислоты из триптофана, всегда имеющегося в корневых выделениях растений.
Процесс внедрения клубеньковых бактерий в ткань корня состоит из двух фаз:
- инфицирование корневых волосков;
- процесс образования клубеньков.
В большинстве случаев внедрившаяся клетка, активно размножается, образует так называемые инфекционные нити и уже в виде таких нитей перемещается в ткани растения. Клубеньковые бактерии, вышедшие из инфекционной нити, продолжают размножаться в ткани хозяина.

Наполняющиеся быстро размножающимися клетками клубеньковых бактерий растительные клетки начинают усиленно делиться. Связь молодого клубенька с корнем бобового растения осуществляется благодаря сосудисто-волокнистым пучкам. В период функционирования клубеньки обычно плотные. К моменту проявления оптимальной активности клубеньки приобретают розовую окраску (благодаря пигменту легоглобину). Фиксировать азот способны лишь те бактерии, которые содержат легоглобин.
Бактерии клубеньков создают десятки и сотни килограммов азотных удобрений на гектаре почвы.
Обмен веществ
Бактерии отличаются друг от друга обменом веществ. У одних он идёт при участии кислорода, у других — без его участия.
Большинство бактерий питается готовыми органическими веществами. Лишь некоторые из них (сине-зелёные, или цианобактерии), способны создавать органические вещества из неорганических. Они сыграли важную роль в накоплении кислорода в атмосфере Земли.
Бактерии впитывают вещества извне, разрывают их молекулы на части, из этих частей собирают свою оболочку и пополняют своё содержимое (так они растут), а ненужные молекулы выбрасывают наружу. Оболочка и мембрана бактерии позволяет ей впитывать только нужные вещества.

Если бы оболочка и мембрана бактерии были полностью непроницаемыми, в клетку не попали бы никакие вещества. Если бы они были проницаемыми для всех веществ, содержимое клетки перемешалось бы со средой — раствором, в которой обитает бактерия. Для выживания бактерии необходима оболочка, которая нужные вещества пропускает, а ненужные — нет.
Бактерия поглощает находящиеся близ неё питательные вещества. Что происходит потом? Если она может самостоятельно передвигаться (двигая жгутик или выталкивая назад слизь), то она перемещается, пока не найдёт необходимые вещества.

Если она двигаться не может, то ждёт, пока диффузия (способность молекул одного вещества проникать в гущу молекул другого вещества) не принесёт к ней необходимые молекулы.
Бактерии в совокупности с другими группами микроорганизмов выполняют огромную химическую работу. Превращая различные соединения, они получают необходимую для их жизнедеятельности энергию и питательные вещества. Процессы обмена веществ, способы добывания энергии и потребности в материалах для построения веществ своего тела у бактерий разнообразны.
Другие бактерии все потребности в углероде, необходимом для синтеза органических веществ тела, удовлетворяют за счёт неорганических соединений. Они называются автотрофами. Автотрофные бактерии способны синтезировать органические вещества из неорганических. Среди них различают:
Хемосинтез
Использование лучистой энергии — важнейший, но не единственный путь создания органического вещества из углекислого газа и воды. Известны бактерии, которые в качестве источника энергии для такого синтеза используют не солнечный свет, а энергию химических связей, происходящих в клетках организмов при окислении некоторых неорганических соединений — сероводорода, серы, аммиака, водорода, азотной кислоты, закисных соединений железа и марганца. Образованное с использованием этой химической энергии органическое вещество они используют для построения клеток своего тела. Поэтому такой процесс называют хемосинтезом.
Важнейшую группу хемосинтезирующих микроорганизмов составляют нитрифицирующие бактерии. Эти бактерии живут в почве и осуществляют окисление аммиака, образовавшегося при гниении органических остатков, до азотной кислоты. Последняя, реагирует с минеральными соединениями почвы, превращаются в соли азотной кислоты. Этот процесс проходит в две фазы.
Железобактерии превращают закисное железо в окисное. Образованная гидроокись железа оседает и образует так называемую болотную железную руду.
Некоторые микроорганизмы существуют за счёт окисления молекулярного водорода, обеспечивая тем самым автотрофный способ питания.
Характерной особенностью водородных бактерий является способность переключаться на гетеротрофный образ жизни при обеспечении их органическими соединениями и отсутствии водорода.
Таким образом, хемоавтотрофы являются типичными автотрофами, так как самостоятельно синтезируют из неорганических веществ необходимые органические соединения, а не берут их в готовом виде от других организмов, как гетеротрофы. От фототрофных растений хемоавтотрофные бактерии отличаются полной независимостью от света как источника энергии.
Бактериальный фотосинтез
Некоторые пигментосодержащие серобактерии (пурпурные, зелёные), содержащие специфические пигменты — бактериохлорофиллы, способны поглощать солнечную энергию, с помощью которой сероводород в их организмах расщепляется и отдаёт атомы водорода для восстановления соответствующих соединений. Этот процесс имеет много общего с фотосинтезом и отличается только тем, что у пурпурных и зелёных бактерий донором водорода является сероводород (изредка — карбоновые кислоты), а у зелёных растений — вода. У тех и других отщепление и перенесение водорода осуществляется благодаря энергии поглощённых солнечных лучей.
Такой бактериальный фотосинтез, который происходит без выделения кислорода, называется фоторедукцией. Фоторедукция углекислого газа связана с перенесением водорода не от воды, а от сероводорода:
6СО 2 +12Н 2 S+hv → С6Н 12 О 6 +12S=6Н 2 О
Биологическое значение хемосинтеза и бактериального фотосинтеза в масштабах планеты относительно невелико. Только хемосинтезирующие бактерии играют существенную роль в процессе круговорота серы в природе. Поглощаясь зелёными растениями в форме солей серной кислоты, сера восстанавливается и входит в состав белковых молекул. Далее при разрушении отмерших растительных и животных остатков гнилостными бактериями сера выделяется в виде сероводорода, который окисляется серобактериями до свободной серы (или серной кислоты), образующий в почве доступные для растения сульфиты. Хемо- и фотоавтотрофные бактерии имеют существенное значение в круговороте азота и серы.
Спорообразование
Внутри бактериальной клетки образуются споры. В процессе спорообразования бактериальная клетка претерпевает ряд биохимических процессов. В ней уменьшается количество свободной воды, снижается ферментативная активность. Это обеспечивает устойчивость спор к неблагоприятным условиям внешней среды (высокой температуре, высокой концентрации солей, высушиванию и др.). Спорообразование свойственно только небольшой группе бактерий.

Споры — не обязательная стадия жизненного цикла бактерий. Спорообразование начинается лишь при недостатке питательных веществ или накоплении продуктов обмена. Бактерии в виде спор могут длительное время находиться в состоянии покоя. Споры бактерий выдерживают продолжительное кипячение и очень длительное проммораживание. При наступлении благоприятных условий спора прорастает и становится жизнеспособной. Спора бактерий — это приспособление к выживанию в неблагоприятных условиях.
Размножение
Размножаются бактерии делением одной клетки на две. Достигнув определённого размера, бактерия делится на две одинаковые бактерии. Затем каждая из них начинает питаться, растёт, делится и так далее.

После удлинения клетки постепенно образуется поперечная перегородка, а затем дочерние клетки расходятся; у многих бактерий в определённых условиях клетки после деления остаются связанными в характерные группы. При этом в зависимости от направления плоскости деления и числа делений возникают разные формы. Размножение почкованием встречается у бактерий как исключение.
При благоприятных условиях деление клеток у многих бактерий происходит через каждые 20-30 минут. При таком быстром размножении потомство одной бактерии за 5 суток способно образовать массу, которой можно заполнить все моря и океаны. Простой подсчёт показывает, что за сутки может образоваться 72 поколения (720 000 000 000 000 000 000 клеток). Если перевести в вес — 4720 тонн. Однако в природе этого не происходит, так как большинство бактерий быстро погибают под действием солнечного света, при высушивании, недостатке пищи, нагревании до 65-100ºС, в результате борьбы между видами и т.д.
Бактерия (1), поглотившая достаточно пищи, увеличивается в размерах (2) и начинает готовиться к размножению (делению клетки). Её ДНК (у бактерии молекула ДНК замкнута в кольцо) удваивается (бактерия производит копию этой молекулы). Обе молекулы ДНК (3,4) оказываются, прикреплены к стенке бактерии и при удлинении бактерии расходятся в стороны (5,6). Сначала делится нуклеотид, затем цитоплазма.

После расхождения двух молекул ДНК на бактерии появляется перетяжка, которая постепенно разделяет тело бактерии на две части, в каждой из которых есть молекула ДНК (7).
Бывает (у сенной палочки), две бактерии слипаются, и между ними образуется перемычка (1,2).

По перемычке ДНК из одной бактерии переправляется в другую (3). Оказавшись в одной бактерии, молекулы ДНК сплетаются, слипаются в некоторых местах (4), после чего обмениваются участками (5).
Роль бактерий в природе
Круговорот
Бактерии — важнейшее звено общего круговорота веществ в природе. Растения создают сложные органические вещества из углекислого газа, воды и минеральных солей почвы. Эти вещества возвращаются в почву с отмершими грибами, растениями и трупами животных. Бактерии разлагают сложные вещества на простые, которые снова используют растения.
Бактерии разрушают сложные органические вещества отмерших растений и трупов животных, выделения живых организмов и разные отбросы. Питаясь этими органическими веществами, сапрофитные бактерии гниения превращают их в перегной. Это своеобразные санитары нашей планеты. Таким образом, бактерии активно участвуют в круговороте веществ в природе.
Почвообразование
Поскольку бактерии распространены практически повсеместно и встречаются в огромном количестве, они во многом определяют различные процессы, происходящие в природе. Осенью опадают листья деревьев и кустарников, отмирают надземные побеги трав, опадают старые ветки, время от времени падают стволы старых деревьев. Всё это постепенно превращается в перегной. В 1 см 3 . поверхностного слоя лесной почвы содержатся сотни миллионов сапрофитных почвенных бактерий нескольких видов. Эти бактерии превращают перегной в различные минеральные вещества, которые могут быть поглощены из почвы корнями растений.
Некоторые почвенные бактерии способны поглощать азот из воздуха, используя его в процессах жизнедеятельности. Эти азотофиксирующие бактерии живут самостоятельно или поселяются в корнях бобовых растений. Проникнув в корни бобовых, эти бактерии вызывают разрастание клеток корней и образование на них клубеньков.
Эти бактерии выделяют азотные соединения, которые используют растения. От растений бактерии получают углеводы и минеральные соли. Таким образом, между бобовым растением и клубеньковыми бактериями существует тесная связь, полезная как одному, так и другому организму. Это явление носит название симбиоза.
Благодаря симбиозу с клубеньковыми бактериями бобовые растения обогащают почву азотом, способствуя повышению урожая.
Распространение в природе
Микроорганизмы распространены повсеместно. Исключение составляют лишь кратеры действующих вулканов и небольшие площадки в эпицентрах взорванных атомных бомб. Ни низкие температуры Антарктики, ни кипящие струи гейзеров, ни насыщенные растворы солей в соляных бассейнах, ни сильная инсоляция горных вершин, ни жёсткое облучение атомных реакторов не мешают существованию и развитию микрофлоры. Все живые существа постоянно взаимодействуют с микроорганизмами, являясь часто не только их хранилищами, но и распространителями. Микроорганизмы — аборигены нашей планеты, активно осваивающие самые невероятные природные субстраты.
Микрофлора почвы
Количество бактерий в почве чрезвычайно велико — сотни миллионов и миллиардов особей в 1 грамме. В почве их значительно больше, чем в воде и воздухе. Общее количество бактерий в почвах меняется. Количество бактерий зависит от типа почв, их состояния, глубины расположения слоёв.
На поверхности почвенных частиц микроорганизмы располагаются небольшими микроколониями (по 20-100 клеток в каждой). Часто они развиваются в толщах сгустков органического вещества, на живых и отмирающих корнях растений, в тонких капиллярах и внутри комочков.
Микрофлора почвы очень разнообразна. Здесь встречаются разные физиологические группы бактерий: бактерии гниения, нитрифицирующие, азотфиксирующие, серобактерии и др. среди них есть аэробы и анаэробы, споровые и не споровые формы. Микрофлора — один из факторов образования почв.
Областью развития микроорганизмов в почве является зона, примыкающая к корням живых растений. Её называют ризосферой, а совокупность микроорганизмов, содержащихся в ней, — ризосферной микрофлорой.
Микрофлора водоёмов
Вода — природная среда, где в большом количестве развиваются микроорганизмы. Основная масса их попадает в воду из почвы. Фактор, определяющий количество бактерий в воде, наличие в ней питательных веществ. Наиболее чистыми являются воды артезианских скважин и родниковые. Очень богаты бактериями открытые водоёмы, реки. Наибольшее количество бактерий находится в поверхностных слоях воды, ближе к берегу. При удалении от берега и увеличении глубины количество бактерий уменьшается.
Чистая вода содержит 100-200 бактерий в 1 мл., а загрязнённая — 100-300 тыс. и более. Много бактерий в донном иле, особенно в поверхностном слое, где бактерии образуют плёнку. В этой плёнке много серо- и железобактерий, которые окисляют сероводород до серной кислоты и тем самым предотвращают замор рыбы. В иле больше спороносных форм, в то время как в воде преобладают неспороносные.
По видовому составу микрофлора воды сходна с микрофлорой почвы, но встречаются и специфические формы. Разрушая различные отбросы, попавшие в воду, микроорганизмы постепенно осуществляют так называемое биологическое очищение воды.
Микрофлора воздуха
Микрофлора воздуха менее многочисленна, чем микрофлора почвы и воды. Бактерии поднимаются в воздух с пылью, некоторое время могут находиться там, а затем оседают на поверхность земли и гибнут от недостатка питания или под действием ультрафиолетовых лучей. Количество микроорганизмов в воздухе зависит от географической зоны, местности, времени года, загрязнённостью пылью и др. каждая пылинка является носителем микроорганизмов. Больше всего бактерий в воздухе над промышленными предприятиями. Воздух сельской местности чище. Наиболее чистый воздух над лесами, горами, снежными пространствами. Верхние слои воздуха содержат меньше микробов. В микрофлоре воздуха много пигментированных и спороносных бактерий, которые более устойчивы, чем другие, к ультрафиолетовым лучам.
Микрофлора организма человека
Тело человека, даже полностью здорового, всегда является носителем микрофлоры. При соприкосновении тела человека с воздухом и почвой на одежде и коже оседают разнообразные микроорганизмы, в том числе и патогенные (палочки столбняка, газовой гангрены и др.). Наиболее часто загрязняются открытые части человеческого тела. На руках обнаруживают кишечные палочки, стафилококки. В ротовой полости насчитывают свыше 100 видов микробов. Рот с его температурой, влажностью, питательными остатками — прекрасная среда для развития микроорганизмов.
Желудок имеет кислую реакцию, поэтому основная масса микроорганизмов в нём гибнет. Начиная с тонкого кишечника реакция становится щелочной, т.е. благоприятной для микробов. В толстых кишках микрофлора очень разнообразна. Каждый взрослый человек выделяет ежедневно с экскрементами около 18 млрд. бактерий, т.е. больше особей, чем людей на земном шаре.
Внутренние органы, не соединяющиеся с внешней средой (мозг, сердце, печень, мочевой пузырь и др.), обычно свободны от микробов. В эти органы микробы попадают только во время болезни.
Бактерии в круговороте веществ
Микроорганизмы вообще и бактерии в частности играют большую роль в биологически важных круговоротах веществ на Земле, осуществляя химические превращения, совершенно недоступные ни растениям, ни животным. Различные этапы круговорота элементов осуществляются организмами разного типа. Существование каждой отдельной группы организмов зависит от химического превращения элементов, осуществляемого другими группами.
Круговорот азота
Циклическое превращение азотистых соединений играет первостепенную роль в снабжении необходимыми формами азота различных по пищевым потребностям организмов биосферы. Свыше 90% общей фиксации азота обусловлено метаболической активностью определённых бактерий.
Круговорот углерода
Биологическое превращение органического углерода в углекислый газ, сопровождающееся восстановлением молекулярного кислорода, требует совместной метаболической активности разнообразных микроорганизмов. Многие аэробные бактерии осуществляют полное окисление органических веществ. В аэробных условиях органические соединения первоначально расщепляются путём сбраживания, а органические конечные продукты брожения окисляются далее в результате анаэробного дыхания, если имеются неорганические акцепторы водорода (нитрат, сульфат или СО 2).
Круговорот серы
Для живых организмов сера доступна в основном в форме растворимых сульфатов или восстановленных органических соединений серы.
Круговорот железа
В некоторых водоёмах с пресной водой содержатся в высоких концентрациях восстановленные соли железа. В таких местах развивается специфическая бактериальная микрофлора — железобактерии, окисляющие восстановленное железо. Они участвуют в образовании болотных железных руд и водных источников, богатых солями железа.
Бактерии являются самыми древними организмами, появившимися около 3,5 млрд. лет назад в архее. Около 2,5 млрд. лет они доминировали на Земле, формируя биосферу, участвовали в образовании кислородной атмосферы.
Бактерии являются одними из наиболее просто устроенных живых организмов (кроме вирусов). Полагают, что они - первые организмы, появившиеся на Земле.
Структурные компоненты бактериальной клетки делят на 2 вида:
- основные структуры (клеточная стенка, цитоплазматическая мембрана с ее производными, цитоплазма с рибосомами и различными включениями, нуклеоид);
- временные структуры (капсула, слизистый чехол, жгутики, ворсинки, эндоспоры, образующиеся лишь на определенных этапах жизненного цикла бактерий).
Основные структуры.
Клеточная стенка находится с внешней стороны от цитоплазматической мембраны. Цитоплазматическая мембрана не входит в состав клеточной стенки. Функции клеточной стенки:
Защита бактерий от осмотического шока и других повреждающих факторов;
Определение формы бактерий;
Участие в метаболизме бактерий.
Клеточная стенка пронизана порами, через которые происходит транспорт экзотоксинов бактерий. Толщина клеточной стенки составляет 10–100 нм. Основной компонент клеточной стенки бактерий - пептидогликан или муреин, состоящий из чередующихся остатков N-ацетил-N-глюкозамина и N-ацетилмурамовой кислоты, соединенных гликозидными связями.
В 1884 году Х. Грам предложил метод окраски бактерий с помощью генцианвиолета, йода, этилового спирта и фуксина. Все бактерии в зависимости от окраски по Граму подразделяют на 2 группы: грамположительные и грамотрицательные бактерии. Клеточная стенка грамположительных бактерий плотно прилегает к цитоплазматической мембране, ее толщина составляет 20-100 нм. В ней имеются тейхоевые кислоты (полимеры глицерина или рибита), а также в небольших количествах полисахариды, белки и липиды. Клеточная стенка грамотрицательных бактерий многослойна, ее толщина составляет 14-17 нм. Внутренний слой (пептидогликан) образует тонкую непрерывную сетку. Внешний слой состоит из фосфолипидов, липопротеина и белков. Белки наружной мембраны прочно связаны с пептидогликановым слоем.
В некоторых условиях бактерии лишаются способности полностью или частично синтезировать компоненты клеточной стенки, в результате чего образуются протопласты, сферопласты и L-формы бактерий. Сферопласты – это бактерии с частично разрушенной клеточной стенкой. Они наблюдаются у грамотрицательных бактерий. Протопласты - это формы, полностью лишенные клеточной стенки. Они образуются грамположительными бактериями. L-формы бактерий - это мутанты бактерий, частично или полностью утратившие способность синтезировать пептидогликан клеточной стенки (бактерии с дефектной клеточной стенкой). Свое название они получили от названия института Листера в Англии, где были открыты в 1935 году.
Цитоплазматическая мембрана (ЦПМ) и ее производные. Цитоплазматическая мембрана (плазмолемма) - это полупроницаемая липопротеидная структура бактериальной клетки, отделяющая цитоплазму от клеточной стенки. Она составляет 8-15% сухой массы клетки. Ее разрушение приводит к гибели клетки. При электронной микроскопии выявлено ее трехслойное строение. Цитоплазматическая мембрана представляет собой комплекс белков (50-75%) и липидов (15-20%). Основная масса липидов представлена фосфолипидами. Кроме того, в составе мембраны обнаружено небольшое количество углеводов.
ЦПМ бактерий выполняет следующие функции:
Барьерная функция (молекулярное “сито”);
Энергетическая;
Избирательный перенос различных органических и неорганических молекул и ионов с помощью специальных переносчиков – транслоказ или пермеаз;
Репликация и последующее разделение хромосомы.
В процессе роста клетки цитоплазматическая мембрана образует многочисленные впячивания (инвагинаты), получившие название мезосом .
Цитоплазма - это содержимое бактериальной клетки, ограниченное цитоплазматической мембраной. Она состоит из цитозоля и структурных элементов.
Цитозоль - гомогенная фракция, включающая растворимые компоненты РНК, ферменты, продукты метаболизма.
Структурные элементы - это рибосомы, внутрицитоплазматические мембраны, включения и нуклеоид.
Рибосомы - органоиды, осуществляющие биосинтез белка. Они состоят из белка и РНК. Представляют собой гранулы диаметром 15-20 нм. Одна бактериальная клетка содержит от 5000 до 50000 рибосом. Рибосомы являются местом синтеза белка.
В цитоплазме прокариотов обнаруживаются различные включения, представляющие запасные вещества клетки. Из полисахаридов в клетках откладываются гликоген, крахмал и крахмалоподобное вещество - гранулеза. Полифосфаты содержатся в гранулах, называемых волютиновыми , или метахроматиновыми , зернами.
Нуклеоид является ядром у прокариотов. Он состоит из одной замкнутой в кольцо двуспиральной нити ДНК, которую рассматривают как бактериальную хромосому. У нуклеоида отсутствует ядерная оболочка.
Кроме нуклеоида в бактериальной клетке обнаружены внехромосомные генетические элементы – плазмиды , которые представляют собой небольшие кольцевые молекулы ДНК, способные к автономной репликации. Роль плазмид состоит в том, что они кодируют дополнительные признаки, дающие клетке преимущества в определенных условиях существования. Наиболее распространены плазмиды, детерминирующие признаки антибиотикорезистентности бактерий (R-плазмиды), синтез энтеротоксинов (Ent-плазмиды) или гемолизинов (Hly-плазмиды).
К временным структурам относятся капсула, жгутики, пили, эндоспоры бактерий.
Капсула - это слизистый слой над клеточной стенкой бактерии. Вещество капсул состоит из нитей полисахаридов. Капсула синтезируется на наружной поверхности цитоплазматической мембраны и выделяется на поверхность клеточной стенки в специфических участках.
Функции капсулы:
Место локализации капсульных антигенов, определяющих вирулентность, антигенную специфичность и иммуногенность бактерий;
Защита клеток от механических повреждений, высыхания, токсических веществ, заражения фагами, действия защитных факторов макроорганизма;
Способность прикрепления клеток к субстрату.
Жгутики – это органы движения бактерий. Жгутики не являются жизненно важными структурами, поэтому могут присутствовать у бактерий или отсутствовать в зависимости от условий выращивания. Количество жгутиков и места их расположения у разных бактерий неодинаково. В зависимости от этого выделяют следующие группы жгутиковых бактерий:
- монотрихи – бактерии с одним полярно расположенным жгутиком;
- амфитрихи – бактерии с двумя полярно расположенными жгутиками или имеющие по пучку жгутиков на обоих концах;
- лофотрихи – бактерии, имеющие пучок жгутиков на одном конце клетки;
- перитрихи – бактерии с множеством жгутиков, расположенных по бокам клетки или на всей ее поверхности.
Химический состав жгутиков представлен белком флагеллином .
К поверхностным структурам бактериальной клетки относятся также ворсинки и пили . Эти структуры участвуют в адсорбции клеток на субстрате (ворсинки, пили общего типа) и в процессах переноса генетического материала (половые пили). Они образованы специфическим гидрофобным белком пилином.
У некоторых бактерий в определенных условиях образуются покоящиеся формы, которые обеспечивают переживание клеток в течение длительного времени в неблагоприятных условиях - эндоспоры . Они устойчивы к неблагоприятным факторам внешней среды.
Расположение спор в клетке:
Центральное (возбудитель сибирской язвы);
Субтерминальное - ближе к концу (возбудитель ботулизма);
Терминальное – на конце палочки (возбудитель столбняка).
Помимо 5 царств живой природы, существует еще два надцарства: прокариоты и эукариоты. Поэтому если рассматривать систематическое положение бактерий, то оно будет следующим:
Почему эти организмы выделяются в отдельный таксон? Все дело в том, что для бактериальной клетки характерно наличие некоторых особенностей, налагающих отпечаток на ее жизнедеятельность и взаимодействие с другими существами и человеком.
Открытие бактерий

Рибосомы - мельчайшие структуры, в большом количестве разбросанные в цитоплазме. Природа их представлена молекулами РНК. Данные гранулы являются материалом, по которому можно определить степень родства и систематическое положение конкретного вида бактерии. Функция их - сборка белковых молекул.
Капсула
Для бактериальной клетки характерно наличие защитных слизистых оболочек, состав которых определяется полисахаридами или полипептидами. Такие структуры имеют название капсул. Различают микро- и макрокапсулы. Данная структура формируется не у всех видов, но у подавляющего большинства, то есть не является обязательной.
От чего защищает капсула бактериальную клетку? От фагоцитоза антителами хозяина, если бактерия патогенная. Либо от высыхания и воздействия вредных веществ, если говорить о других видах.
Слизь и включения
Также необязательные структуры бактерий. Слизь, или гликокаликс, по химической основе является мукоидным полисахаридом. Может формироваться как внутри клетки, так и наружными ферментами. Хорошо растворима в воде. Предназначение: прикрепление бактерии к субстрату - адгезия.
Включения - это микрогранулы в цитоплазме различной химической природы. Это могут быть белки, аминокислоты, нуклеиновые кислоты или полисахариды.

Органоиды движения
Особенности бактериальной клетки также проявляются и в ее движении. Для этого присутствуют жгутики, которые могут быть в разном количестве (от одного до нескольких сотен на клетку). Основа каждого жгутика - белок флагеллин. Благодаря эластичным сокращениям и ритмичным движениям из стороны в сторону бактерия может передвигаться в пространстве. Крепится жгутик к цитоплазматической мембране. Расположение также может варьироваться у разных видов.
Пили
Еще более тонкие, чем жгутики, структуры, принимающие участие в:
- прикреплении к субстрату;
- водно-солевом питании;
- половом размножении.
Состоят из белка пилина, количество их может доходить до нескольких сотен на клетку.
Сходство с клетками растений
Бактериальная и имеют одно неоспоримое сходство - наличие клеточной стенки. Однако если у растений она есть бесспорно, то у бактерий присутствует не у всех видов, то есть относится к необязательным структурам.
Химический состав бактериальной клеточной стенки:
- пептидогликан муреин;
- полисахариды;
- липиды;
- белки.
Обычно данная структура имеет двойной слой: наружный и внутренний. Функции выполняет такие же, как растений. Поддерживает и обозначает постоянную форму тела и обеспечивает механическую защиту.

Образование спор
Каково строение бактериальной клетки, мы рассмотрели достаточно подробно. Осталось только упомянуть о том, как бактерии могут переживать неблагоприятные условия, очень долгое время не теряя жизнеспособности.
Это им удается путем формирования структуры под названием спора. Она не имеет отношения к размножению и лишь предохраняет бактерии от неблагоприятных условий. По форме споры могут быть различными. При восстановлении нормальных окружающих условий спора инициируется и прорастает в активную бактерию.
Клетка прокариотических организмов имеет сложное строго упорядоченное строение и обладает принципиальными особенностями ультраструктурной организации и химического состава.
Структурные компоненты бактериальной клетки делят на основные и временные (рис. 2). Основными структурами являются: клеточная стенка, цитоплазматическая мембрана с ее производными, цитоплазма с рибосомами и различными включениями, нуклеоид; временные — капсула, слизистый чехол, жгутики, ворсинки, эндоспоры, образующиеся лишь на определенных этапах жизненного цикла бактерий, у некоторых видов они отсутствуют полностью.
У прокариотической клетки структуры, расположенные снаружи от цитоплазматической мембраны, называют поверхностными (клеточная стенка, капсула, жгутики, ворсинки).
Термин «оболочка» в настоящее время используется для обозначения клеточной стенки и капсулы бактерий или только клеточной стенки, цитоплазматическая мембрана не входит в состав оболочки и относится к протопласту.
Клеточная стенка — важный структурный элемент бактериальной клетки, располагающийся между цитоплазматической мембраной и капсулой; у бескапсульных бактерий — это внешняя оболочка клетки. Она обязательна для всех прокариот, за исключением микоплазм и L-форм бактерий. Выполняет ряд функций: защищает бактерии от осмотического шока и других повреждающих факторов, определяет их форму, участвует в метаболизме; у многих видов патогенных бактерий токсична, содержит поверхностные антигены, а также несет на поверхности специфические рецепторы для фагов. В клеточной стенке бактерий имеются поры, которые участвуют в транспорте экзотоксинов и других экзобелков бактерий. Толщина клеточной стенки 10—100 нм, и на ее долю приходится от 5 до 50 % сухих веществ клетки.
Основным компонентом клеточной стенки бактерий является пептидогликан, или муреин (лат. murus — стенка), — опорный полимер, имеющий сетчатую структуру и образующий ригидный (жесткий) наружный каркас бактериальной клетки. Пептидогликан имеет основную цепь (остов), состоящую из чередующихся остатков N-ацстил-М-глюкозамина и N-ацетилмурамовой кислоты, соединенных 1,4-гликозидными связями, идентичные тетрапептидные боковые цепочки, прикрепляющиеся к молекулам N-ацстилмурамовой кислоты, и короткие поперечные пептидные мостики, связывающие полисахаридные цепи. Два типа связей (гликозидные и пептидные), которые соединяют субъединицы пептидогликана, придают этому гетерополимеру структуру молекулярной сети. Остов пептидогликанового слоя у всех видов бактерий одинаков; тетрапептидные белковые цепочки и пептидные (поперечные) у неодинаковых видов различны.
По тинкториальным свойствам все бактерии подразделяются на две группы: грамположительные и грамотрицателъные. В 1884 г. X. Грам предложил метод окраски, который был использован для дифференцирования бактерий. Сущность метода состоит в том, что грамположительные бактерии прочно фиксируют комплекс генцианвиолета и йода, не подвергаются обесцвечиванию этанолом и поэтому не воспринимают дополнительный краситель фуксин, оставаясь окрашенными в фиолетовый цвет. У грамотрицательных бактерий этот комплекс легко вымывается из клетки этанолом, и они при дополнительном нанесении фуксина окрашиваются в красный цвет. У некоторых бактерий положительная окраска по Граму наблюдается только в стадии активного роста. Способность прокариот окрашиваться по методу Грама или обесцвечиваться этанолом определяется спецификой химического состава и ультраструктуры их клеточной стенки. Пептидогликан у грамположительных бактерий — основной компонент клеточной стенки и составляет от 50 до 90 %, у грамотрицательных — 1 —10 %. Структурные микрофибриллы пептидогликана грамотрицательных бактерий сшиты менее компактно, поэтому поры в их пептидогликановом слое значительно шире, чем в молекулярном каркасе грамположитсльных бактерий. При такой структурной организации пептидогликана фиолетовый комплекс генцианвиолета и йода у грамотрицательных бактерий будет вымываться быстрее.
Клеточная стенка грамположительных бактерий плотно прилегает к цитоплазматической мембране, массивна, се толщина находится в пределах 20—100 нм. Для нее характерно наличие тейхоевых кислот, они связаны с пептидогликаном и представляют собой полимеры трехатомного спирта — глицерина или пятиатомного спирта — рибита, остатки которых соединены фосфодиэфирными связями. Тейхоевые кислоты связывают ионы магния и участвуют в транспорте их в клетку. В составе клеточной стенки грамположительных прокариот в небольших количествах также найдены полисахариды, белки и липиды.
Рис. 2. Схема строения прокариотической клетки:
1 — капсула; 2 — клеточная стенка; 3 — цитоплазматическая мембрана; 4 — нуклеоид; 5 — цитоплазма; 6 — хроматофоры; 7 — тилакоиды; 8 — мезосома; 9 — рибосомы; 10 — жгутики; 11— базальное тельце; 12 — пили; 13 — включение серы; 14 — капли жира; 15 — гранулы полифосфата; 16 — плазмида
Клеточная стенка грамотрицательных бактерий многослойна, толщина ее 14—17 нм. Внутренний слой — пептидогликан, который образует тонкую (2 нм) непрерывную сетку, окружающую клетку. Пептидогликан содержит только мезодиаминопимелиновую кислоту и не имеет лизина. Внешний слой клеточной стенки — наружная мембрана — состоит из фосфолипидов, липополисахарида, липопротеина и белков. В наружной мембране содержатся белки основы (матричные), они прочно связаны с пептидогликановым слоем. Одной из их функций является формирование в мембране гидрофильных пор, через которые осуществляется диффузия молекул с массой до 600, иногда 900. Матричные белки, кроме того, выполняют еще роль рецепторов для некоторых фагов. Липополисахарид (ЛПС) клеточных стенок грамотрицательных бактерий состоит из липида А и полисахарида. Токсичный для животных ЛПС получил название эндотоксина. Тейхоевые кислоты у грамотрицательных бактерий не обнаружены.
Структурные компоненты клеточной стенки грамотрицальных бактерий отграничены от цитоплазматической мембраны и разделены промежутком, называемым периплазмой или периплазматическим пространством.
Протопласты и сферопласты. Протопласты — формы прокариот, полностью лишенные клеточной стенки, образующиеся обычно у грамположительных бактерий. Сферопласты — бактерии с частично разрушенной клеточной стенкой. У них сохраняются элементы наружной мембраны. Наблюдаются у грамотрицательных бактерий и значительно реже у грамположительных. Образуются в результате разрушения пептидогликанового слоя литическими ферментами, например лизоцимом, или блокирования биосинтеза пептидогликана антибиотиком пенициллином и др. в среде с соответствующим осмотическим давлением.
Протопласты и сферопласты имеют сферическую или полусферическую форму и в 3—10 раз крупнее исходных клеток. В обычных условиях наступает осмотический лизис и они погибают. В условиях повышенного осмотического давления способны некоторое время переживать, расти и даже делиться. При снятии фактора, разрушающего пептидогликан, протопласты, как правило, отмирают, но могут превращаться в L-формы; сферопласты легко реверсируют в исходные бактерии, иногда трансформируются в L-формы или же гибнут.
L-Формы бактерий. Это фенотипические модификации, или мутанты, бактерий, частично или полностью утратившие способность синтезировать пептидогликан клеточной стенки. Таким образом, L-формы — бактерии, дефектные по клеточной стенке. Свое название они получили в связи с тем, что были выделены и описаны в институте Листера в Англии в 1935 г. Образуются при воздействии L-трансформирующих агентов — антибиотиков (пенициллина, полимиксина, бацитрацина, венкомицина, стрептомицина), аминокислот (глицина, метионина, лейцина и др.), фермента лизоцима, ультрафиолетовых и рентгеновых лучей. В отличие от протопластов и сферопластов L-формы обладают относительно высокой жизнеспособностью и выраженной способностью к репродукции. По морфологическим и культуральным свойствам они резко отличаются от исходных бактерий, что обусловлено утратой клеточной стенки и изменением метаболической активности.
L-Формы бактерий полиморфны. Встречаются элементарные тельца размером 0,2—1 мкм (минимальные репродуцирующие элементы), шары — 1—5, большие тела — 5—50, нити — до 4 мкм и более. Клетки L-форм имеют хорошо развитую систему внутрицитоплазматических мембран и миелиноподобные структуры. Вследствие дефекта клеточной стенки осмотически неустойчивы и их можно культивировать только на специальных средах с высоким осмотическим давлением; они проходят через бактериальные фильтры.
Различают стабильные и нестабильные L-формы бактерий. Первые полностью лишены ригидной клеточной стенки, что сближает их с протопластами; они крайне редко реверсируют в исходные бактериальные формы. Вторые могут обладать элементами клеточной стенки, в чем они проявляют сходство со сферопластами; в отсутствие фактора, вызвавшего их образование, реверсируют в исходные клетки.
Процесс образования L-форм получил название L-трансформации или L-индукции. Способностью к L-трансформации обладают практически все виды бактерий, в том числе и патогенные (возбудители бруцеллеза, туберкулеза, листерии и др.).
L-Формам придается большое значение в развитии хронических рецидивирующих инфекций, носительстве возбудителей, длительной персистенции их в организме. Доказана трансплацентарная инвазивность элементарных телец L-форм бактерий.
Инфекционный процесс, вызванный L-формами бактерий, характеризуется атипичностью, длительностью течения, тяжестью заболевания, трудно поддается химиотерапии.
Капсула — слизистый слой, расположенный над клеточной стенкой бактерии. Вещество капсулы четко отграничено от окружающей среды. В зависимости от толщины слоя и прочности соединения с бактериальной клеткой различают макрокапсулу, толщиной более 0,2 мкм, хорошо различимую в световом микроскопе, и микрокапсулу, толщиной менее 0,2 мкм, обнаруживаемую лишь при помощи электронного микроскопа или выявляемую химическими и иммунологическими методами. Макрокапсулу (истинную капсулу) образуют В. anlhracis, C1. perfringens, микрокапсулу — Escherichia coJi. Капсула не является обязательной структурой бактериальной клетки: потеря ее не приводит к гибели бактерии. Известны бескапсульные мутанты бактерий, например сибиреязвенный вакцинный штамм СТИ-1.
Вещество капсул состоит из высокогидрофильных мицелл, химический же состав их весьма разнообразен. Основные компоненты большинства капсул прокариот — гомо- или гетсрополисахариды (энтсробактерии и др.). У некоторых видов бацилл капсулы построены из полипептида. Так, в состав капсулы В. anthracis входит полипептид Д-глутаминовой кислоты (правовращающий изомер). В состав микрокапсулы микобактерий туберкулеза млекопитающих входят гликопептиды, представленные сложным эфиром трегалозы и миколовой кислоты (корд-фактор).
Синтез капсулы — сложный процесс и у различных прокариот имеет свои особенности; считают, что биополимеры капсулы синтезируются на наружной поверхности цитоплазматической мембраны и выделяются на поверхность клеточной стенки в определенных специфических ее участках.
Существуют бактерии, синтезирующие слизь, которая откладывается на поверхности клеточной стенки в виде бесструктурного слоя полисахаридной природы. Слизистое вещество, окружающее клетку, по толщине часто превосходит диаметр последней. У сапрофитной бактерии лейконостока наблюдается образование одной капсулы для многих особей. Такие скопления бактерий, заключенных в общую капсулу, называются зооглеями.
Капсула — полифункциональный органоид, выполняющий важную биологическую роль. Она является местом локализации капсульных антигенов, определяющих вирулентность, антигенную специфичность и иммуногенность бактерий. Утрата капсулы у патогенных бактерий резко снижает их вирулентность, например у бескапсульных штаммов бациллы антракса. Капсулы обеспечивают выживание бактерий, защищая их от механических повреждений, высыхания, заражения фагами, токсических веществ, а у патогенных форм — от действия защитных сил макроорганизма: инкапсулированные клетки плохо фагоцитируются. У некоторых видов бактерий, в том числе и патогенных, способствует прикреплению клеток к субстрату.
В ветеринарной микробиологии выявление капсулы используют в качестве дифференциального морфологического признака возбудителя при исследовании на сибирскую язву.
Для окрашивания капсул применяют специальные методы — Романовского — Гимзы, Гинса — Бурри, Ольта, Михина и др.
Микрокапсулу и слизистый слой определяют серологическими реакциями (РА), антигенные компоненты капсулы идентифицируют при помощи иммунофлюоресцентного метода (РИФ) и РДД.
Жгутики — органоиды движения бактерий, представленные тонкими, длинными, нитевидными структурами белковой природы. Их длина превышает бактериальную клетку в несколько раз и составляет 10—20 мкм, а у некоторых спирилл достигает 80— 90 мкм. Нить жгутика (фибрилла) — полный спиральный цилиндр диаметром 12—20 нм. У вибрионов и протея нить окружена футляром толщиной 35 нм.
Жгутик состоит из трех частей: спиральной нити, крюка и базального тельца. Крюк — изогнутый белковый цилиндр, выполняющий функцию гибкого связывающего звена между базальным тельцем и жесткой нитью жгутика. Базальное тельце — сложная структура, состоящая из центрального стержня (оси) и колец.

Рис. 3. Жгутики:
а — монотрихи; б — амфитрихи; в — лофотрихи; г — перитрихи
Жгутики не являются жизненно важными структурами бактериальной клетки: существуют фазовые вариации бактерий, когда в одной фазе развития клетки они имеются, у другой — отсутствуют. Так, у возбудителя столбняка в старых культурах преобладают клетки без жгутиков.
Количество жгутиков (от I до 50 и более) и места их локализации у бактерий разных видов неодинаковы, но стабильны для одного вида. В зависимости от этого выделяют следующие группы жгутиковых бактерий: моиотрихи — бактерии с одним полярно расположенным жгутиком; амфитрихи — бактерии с двумя полярно расположенными жгутиками или имеющие по пучку жгутиков на обоих концах; лофотрихи — бактерии, имеющие пучок жгутиков на одном конце клетки; перитрихи — бактерии с множеством жгутиков, расположенных по бокам клетки или на всей ее поверхности (рис. 3). Бактерии, не имеющие жгутиков, называют атрихиями.
Будучи органами движения, жгутики типичны для плавающих палочковидных и извитых форм бактерий и лишь в единичных случаях встречаются у кокков. Они обеспечивают эффективное движение в жидкой среде и более медленное перемещение по поверхности твердых субстратов. Скорость движения монотрихов и лофотрихов достигает 50 мкм/с, амфитрихи и перитрихи движутся медленнее и обычно за 1 с проходят расстояние, равное размерам их клетки.
Бактерии передвигаются беспорядочно, однако они способны к направленным формам движения — таксисам, которые определяются внешними стимулами. Реагируя на различные факторы окружающей среды, бактерии за короткое время локализуются в оптимальной зоне обитания. Таксис может быть положительным и отрицательным. Принято различать: хемотаксис, аэротаксис, фототаксис, магнототаксис. Хемотаксис вызывается разницей в концентрации химических веществ в среде, аэротаксис — кислорода, фототаксис — интенсивностью освещения, магнитотаксис определяется способностью микроорганизмов ориентироваться в магнитном поле.
Выявление подвижных жгутиковых форм бактерий имеет значение для их идентификации при лабораторной диагностике инфекционных болезней.
Пили (фимбрии, ворсинки) — прямые, тонкие, полые белковые цилиндры толщиной 3—25 нм и длиной до 12 мкм, отходящие от поверхности бактериальной клетки. Образованы специфическим белком — пилином, берут начало от цитоплазматической мембраны, встречаются у подвижных и неподвижных форм бактерий и видимы только в электронном микроскопе (рис. 4). На поверхности клетки может быть от 1—2, 50—400 и более пилей до нескольких тысяч.

Рис. 4. Пили
Существует два класса пилей: половые (секспили) и пили общего типа, которые чаще называют фимбриями. У одной и той же бактерии могут быть пили разной природы. Половые пили возникают на поверхности бактерий в процессе конъюгации и выполняют функцию органелл, через которые происходит передача генетического материала (ДНК) от донора к реципиенту.
Пили общего типа располагаются перитрихиально (кишечная палочка) или на полюсах (псевдомонады); одна бактерия их может содержать сотни. Они принимают участие в слипании бактерий в агломераты, прикреплении микробов к различным субстратам, в том числе к клеткам (адгезивная функция), в транспорте метаболитов, а также способствуют образованию пленок на поверхности жидких сред; вызывают агглютинацию эритроцитов.
Цитоплазматическая мебрана и ее производные. Цитоплазматическая мембрана (плазмолемма) — полупроницаемая липопротеидная структура бактериальных клеток, отделяющая цитоплазму от клеточной стенки. Она является обязательным полифункциональным компонентом клетки и составляет 8—15 % ее сухой массы. Разрушение цитоплазматической мембраны приводит к гибели бактериальной клетки. На ультратонких срезах в электронном микроскопе выявляется ее трехслойное строение — два ограничивающих осмиофильных слоя, толщиной 2—3 нм каждый, и один осмиофобный центральный слой толщиной 4—5 нм.
Цитоплазматическая мембрана в химическом отношении — белково-липидный комплекс, состоящий из 50—75 % белков и 15—50 % липидов. Основная часть мембранных липидов (70— 90 %) представлена фосфолипидами. Она построена из двух мономолекулярных белковых слоев, между которыми расположен липидный слой, состоящий из двух рядов правильно ориентированных молекул липидов.
Цитоплазматичсская мембрана служит осмотическим барьером клетки, контролирует поступление питательных веществ в клетку и выход продуктов метаболизма наружу, в ней содержатся субстратспецифические ферменты-пермеазы, осуществляющие активный избирательный перенос органических и неорганических молекул.
Ферменты цитоплазматической мембраны катализуют конечные этапы синтеза мембранных липидов, компонентов клеточной стенки, капсулы и экзоферментов; на мембране локализованы ферменты окислительного фосфорилирования и ферменты транспорта электронов, ответственные за синтез энергии.
В процессе роста клетки цитоплазмзтическая мембрана образует многочисленные инвагинаты, формирующие внутрицитоплазмати-ческие мембраны структуры. Локальные инвагинаты мембраны получили название мезосом. Эти структуры хорошо выражены у грамположительных бактерий, хуже — у грамотрицательных и плохо — у риккетсий и микоплазм.
Установлена связь мезосом с хромосомой бактерии, такие структуры называются нуклеоидосомеши. Интегрированные с нуклеоидом мезосомы принимают участие в кариокинезе и цитокинезе микробных клеток, обеспечивая распределение генома после окончания репликации ДНК и последующее расхождение дочерних хромосом. Мезосомы, как и цитоплазматическая мембрана, являются центрами дыхательной активности бактерий, поэтому их иногда называют аналогами митохондрий. Однако значение мезосом окончательно еще не выяснено. Они увеличивают рабочую поверхность мембран, возможно, выполняют только структурную функцию, производя разделение бактериальной клетки на относительно обособленные отсеки, что создает более благоприятные условия для протекания ферментативных процессов. У патогенных бактерий обеспечивают транспорт белковых молекул экзотоксинов.
Цитоплазма — содержимое бактериальной клетки, отграниченное цитоплазматической мембраной. Состоит из цитозоля — гомогенной фракции, включающей растворимые компоненты РНК, вещества субстрата, ферменты, продукты метаболизма, и структурных элементов — рибосом, внутрицитоплазматических мембран, включений и нуклеоида.
Рибосомы — органоиды, осуществляющие биосинтез белка. Состоят из белка и РНК, соединенных в комплекс водородными и гидрофобными связями. Бактериальные рибосомы — гранулы диаметром 15—20 нм, имеют константу седиментации 70S и образованы из двух рибонуклеопротеидных субъединиц: 30S и 50S. Одна бактериальная клетка может содержать от 5000—50 000 рибосом, посредством и-РНК они объединяются в полисомы-агрегаты, состоящие из 50—55 рибосом, обладающих высокой белоксинтезирующей активностью.
В цитоплазме бактерий выявляются различного типа включения. Они могут быть твердыми, жидкими и газообразными, с белковой мембраной или без нее и присутствовать непостоянно. Значительная часть их представляет собой запасные питательные вещества и продукты клеточного метаболизма. К запасным питательным веществам относятся: полисахариды, липиды, полифосфаты, отложения серы и др. Из включений полисахаридной природы чаще обнаруживаются гликоген и крахмалоподобное вещество гранулеза, которые служат источником углерода и энергетическим материалом. Липиды накапливаются в клетках в виде гранул и капелек жира, к ним относятся окруженные мембраной гранулы поли-/3-оксимас-ляной кислоты, резко преломляющие свет и хорошо различимые в световом микроскопе. Выявляются и бациллы антракса и аэробных спорообразующих сапрофитных бактерий. Микобактерии в качестве запасных веществ накапливают воски. В клетках некоторых кори-небактерий, спирилл и других содержатся гранулы волютина, образованные полифосфатами. Они характеризуются метахромазией: толуидиновый синий и метиленовый синий окрашивают их в фиолетово-красный цвет. Волютиновые гранулы играют роль фосфатных депо.
К включениям, окруженным мембраной, также относятся газовые вакуоли, или аэросомы, они снижают удельную массу клеток, встречаются у водных прокариот.
Нуклеоид — ядро у прокариот. Он состоит из одной замкнутой в кольцо двухспиральной нити ДНК длиной 1,1 —1,6 нм, которую рассматривают как одиночную бактериальную хромосому, или генофор.
Нуклеоид у прокариот не отграничен от остальной части клетки мембраной — у него отсутствует ядерная оболочка.
В состав структур нуклеоида входят РНК-полимераза, основные белки и отсутствуют гистоны; хромосома закрепляется на цитоплазматической мембране, а у грамположительных бактерий — на мезосомс. Бактериальная хромосома реплицируется поликонсервативным способом: родительская двойная спираль ДНК раскручивается и на матрице каждой полинуклеотидной цепи собирается новая комплементарная цепочка. Нуклеоид не имеет митотического аппарата, и расхождение дочерних ядер обеспечивается ростом цитоплазматической мембраны.
Бактериальное ядро — дифференцированная структура. В зависимости от стадии развития клетки нуклеоид может быть дискретным (прерывистым) и состоять из отдельных фрагментов. Это связано с тем, что деление бактериальной клетки во времени осуществляется после завершения цикла репликации молекулы ДНК и оформления дочерних хромосом.
В нуклеоиде сосредоточен основной объем генетической информации бактериальной клетки.
Кроме нуклеоида в клетках многих бактерий обнаружены внехромосомные генетические элементы — плазмиды, представленные небольшими кольцевыми молекулами ДНК, способными к автономной репликации.