Термин «рост» в применении к микроорганизмам означает увеличение размеров отдельной особи, а «размножение» — повышение числа особей в популяции. При росте микробной клетки объем ее возрастает значительно быстрее, чем поверхность, поэтому распределение питательных веществ в цитоплазме клетки становится менее эффективным и клетка делится. Перед делением ее происходит удвоение молекул ДНК. Каждая дочерняя клетка получает копию материнской ДНК.
Быстрота размножения разных микробов, выращиваемых в одинаковых условиях, различна. Для большинства бактерий период генерации (время, прошедшее роорганизмы могут использовать большой набор окисляемых органических соединений, чаще всего глюкозу. Энергия получается из этих соединений в результате их окисления или, точнее, отдачи ими электронов.
Совокупность биохимических процессов, в результате которых освобождается энергия, необходимая для жизнедеятельности клетки, называется дыханием, или биологическим окислением. Применительно к микроорганизмам говорят об анаэробном и аэробном типах дыхания.
Между двумя последовательными делениями клетки) в среднем равен 15—30 мин; например, для кишечной палочки —15—17 мин, возбудителей брюшного тифа — 23 мин, коринебактерий дифтерии — 34 мин. Микобактерии туберкулеза делятся медленнее — один раз за 18 ч, спирохеты — за 10 ч.
Способы размножения у различных групп микроорганизмов неодинаковы: бактерии, риккетсии, спирохеты размножаются путем поперечного деления на две равноценные особи. Грамположительные бактерии делятся путем образования перегородки, врастающей от периферии к центру. У микобактерий туберкулеза поперечная перегородка образуется внутри клетки, затем она расщепляется на два слоя и клетка делится на две части, В образовании перегородки принимает участие как цитоплазматическая мембрана, так и клеточная стенка. По-видимому, в процессе деления бактерий активное участие принимает мезосома, тесно связанная с цитоплазматической мембраной. Грамотрицательные бактерии и риккетсии истончаются в центре и делятся перетяжкой на две особи. Размножение клубеньковых бактерий и фраициселл происходит путем образования почки, которая, по величине меньше исходной клетки. У бактерий существует также процесс конъюгации — временного соединения двух особей.
Рост бактерий и спирохет не всегда сопровождается их делением. Соли желчных кислот, мыла, пенициллин, ультрафиолетовые лучи задерживают деление клетки, в результате чего образуются длинные нити значительно большего размера, чем исходные клетки.
При внесении бактерий в питательную среду различают фазы их роста и размножения, которые определяются наличием доступных источников питания и накоплением токсических продуктов обмена (рис. 21).
Первая фаза — латентная (лаг-фаза) — соответствует приспособлению бактерий к новым условиям существования. В этот период бактерии адаптируются к питательной среде, роста их не наблюдается.
Вторая фаза — логарифмического роста (экспоненциальная), когда бактерии энергично растут, увеличиваются, при достижении определенного размера начинают делиться на две дочерние клетки. Деление в этот период происходит с постоянной скоростью. Среднее время генерации (или удвоения) для каждого вида бактерий различно. В это время бактерии извлекают из среды питательные вещества, в результате чего в ней начинают накапливаться продукты обмена.
Третья фаза — стационарного роста, во время которой число организмов в культуре все время остается постоянным. В этот период в питательной среде количество питательных веществ значительно уменьшается, а накопление продуктов обмена увеличивается. Условия жизни для микроорганизмов становятся все менее благоприятными. Длительность стационарной фазы у разных бактерий различная.
Четвертая фаза — отмирания, когда клеток бактерий становится все меньше и они погибают. В конце этой фазы число отмирающих бактерий начинает преобладать над числом жизнеспособных клеток. Полная гибель микробов в культуре может наступить через несколько недель или месяцев, что зависит от вида микроба, реакции среды и других факторов.
Простейшие могут размножаться поперечным делением, перетяжкой на две равноценные особи — амебы и продольным делением — трипаносомы, лямблии, балантидии. Балантидии перед делением на две особи могут обмениваться своими ядрами — микронуклеусами (процесс конъюгации), малярийный плазмодий имеет бесполый и половой цикл развития.
Вирусы размножаются (репродуцируются) только внутри живой клетки организма хозяина.
Процесс воспроизведения вируса складывается из нескольких этапов:
1) проникновение вируса в клетку;
2) внутриклеточное размножение;
3) созревание вируса и образование внешних оболочек у некоторых вирусов; 4) выделение вируса из клетки.
Процесс проникновения вируса в чувствительную клетку начинается с его адсорбции на поверхности клетки, обладающей специфическими вирусными рецепторами. Процесс освобождения нуклеиновой кислоты от капсида и внешних оболочек начинается уже в цитоплазматической мембране клетки и заканчивается в цитоплазме (вирус гриппа, осповакцины).
Фаза внутриклеточного размножения вируса, или его воспроизведение, начинается обычно с процессов подавления клеточного макромолекулярного синтеза. Все энергетические системы клетки, ее ферменты, РНК, рибосомы начинают работать на воспроизведение вируса. Пораженная клетка поставляет вирусу нуклеотиды для построения нуклеиновых кислот, аминокислоты — для белков. Репликация (англ. replicate — копировать, повторять) вирусной РНК осуществляется с помощью ферментов — полимераз, а матрицей служит сама молекула РНК вируса. У ДНК-содержащих вирусов на матрице ДНК в ядре клетки синтезируется специфическая РНК, которая затем определяет синтез вирусной ДНК и белка. Белки вирусов синтезируются в рибосомах клетки.
Созревание вирусной частицы, заключение вирусной нуклеиновой кислоты в капсид, происходит в ядре пораженной клетки (герпесвирусы, аденовирусы) или в цитоплазме (вирусы группы оспы, рабдовирусы, пикорнавирусы). Формирование внешних оболочек у миксовирусов, тогавирусов происходит при прохождении через цитоплазматическую мембрану клетки хозяина. Вирус герпеса часть своей внешней оболочки получает, проходя через мембрану ядра клетки.
Выделение вируса из клетки может происходить по-разному. Миксовирусы и тогавирусы по мере созревания могут часами выделяться клеткой без ее повреждения. Вирус полиомиелита (не имеющий внешней оболочки) образуется в клетке быстро, остается в ней долго и выделяется мгновенно, в виде вспышки. Конечным результатом взаимодействия вируса и клетки хозяина могут быть быстрая деструкция и гибель клетки. Иногда вирусы могут долго присутствовать в клетке, не вызывая ее гибели, и сохраняются в бесконечном числе клеточных генераций — латентные вирусы. В некоторых случаях вирус может разрушаться клеткой без видимых последствий для нее (абортивная вирусная инфекция).
Рост и размножение бактерий
Рост бактерий происходит в результате множества взаимосвязанных биохимических реакций, осуществляющих синтез клеточного материала, что приводит к увеличению количества всех химических компонентов. У бактерий различают индивидуальный рост бактериальной клетки и рост бактерий в популяции.
Индивидуальный рост бактерий . О нем судят по увеличению размеров отдельных особей. Скорость роста зависит от внешних условий и физиологического состояния самой клетки. При постоянных условиях рост осуществляется с постоянной скоростью. Палочковидные бактерии растут преимущественно в направлении длинной оси, поэтому соотношение между поверхностью клетки и ее объемом при росте клеток существенно не изменяется, и это создает постоянные условия снабжения каждой части клетки питательными веществами и кислородом. Кокки растут равномерно во всех направлениях, увеличивая размеры радиуса клетки, при этом относительная величина поверхности клетки падает, поэтому условия снабжения каждой части клетки становятся все более неблагоприятными. В промежутках между клеточными делениями бактерии имеют большие размеры, чем сразу после деления.
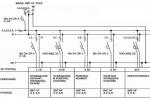
Размножение бактерий. Наиболее часто бактерии размножаются путем бинарного деления, когда из одной клетки образуется две, каждая из которых вновь делится. Процессу деления всегда предшествует репликация ДНК. Существует два типа деления – деление перетяжкой (перешнуровывание) и с помощью поперечной перегородки (рисунок 1.9).

Рисунок 1.9 – Деление бактерий
А - деление перетяжкой; Б - деление поперечной перегородкой; КС – клеточная стенка; ЦМ – цитоплазматическая мембрана; Н – нуклеоид; П – перетяжка
Деление перетяжкой (констрикция) сопровождается сужением клетки в месте ее деления, и в этом процессе принимают участие все слои клеточных оболочек. Выпячивание оболочек с обеих сторон внутрь клетки все более ее сужает и, наконец, делит на две. Так делятся многие грамотрицательным бактериям.
Деление с образованием поперечной перегородки присуще грамположительным бактериям. Однако у некоторых групп бактерий отмечена смена способов деления (тионовые бактерии, микобактерии). У шаровидных бактерий может образовываться несколько поперечных перегородок (тетракокки, сарцины).
Период от деления до деления называется клеточным циклом (онтогенез бактерий). Различают несколько типов вегетативного клеточного цикла: мономорфный – образуется только один морфологический тип клеток (например, бациллы), диморфный – два морфологических типа, полиморфный – несколько, каждый из которых характеризуется определенными и постоянными особенностями клеточного цикла (например, актиномицеты). При диморфном и полиморфном циклах различают дочерние и материнские клетки.
Почкование убактерий является разновидностью бинарного деления. Этот способ размножения присущ бактериям, имеющим диморфные или полиморфные клеточные циклы. Почкующимся бактериям присуща полярность клеток. Некоторые бактерии размножаются с помощью экзоспор (но не эндоспор!), фрагментами гиф (актиномицеты). Есть бактерии, у которых имеются половые ворсинки, или F-пили (англ. fertility –фертильность, плодовитость), обусловленные наличием полового фактора.
Бактерии характеризуются высокой скоростью размножения. Например, при благоприятных условиях кишечная палочка делится каждые 20…30 мин, за сутки из одной клетки проучится 2 72 (72 поколения). В условиях, исключающих гибель, такая биомасса составит 4720 т. Скорость размножения зависит от факторов внешней среды (температура, условия питания, влажность, реакция среды и др.) и от видовых особенностей бактерий. Высокая скорость размножения бактерий обеспечивает их сохранение на Земле даже в условиях массовой гибели. Сохранившиеся отдельные клетки размножаются и вновь дают поколение.
Рост бактерий в популяции. Популяция (фр. population – население) – это совокупность бактерий одного вида (чистая культура) или разных видов (смешанная ассоциация), развивающихся в ограниченном пространстве (например, в питательной среде). В бактериальной популяции постоянно происходит рост, размножение и отмирание клеток. Культивирование микроорганизмов в искусственных условиях бывает периодическим, непрерывным и синхронным.
Периодическое (стационарное) культивирование . Это культивирование происходит без притока и оттока питательной среды. Оно характеризуется классической кривой роста микроорганизмов, в которой выделяют отдельные фазы роста бактериальной популяции, отражающие общую закономерность роста и размножения клеток (рисунок 1.10).

Рисунок 1.10 – Кривая роста и развития бактериальной популяции
Лаг-фаза (англ. lag – отставание) начинается с момента посева бактерий в свежую питательную среду. Клетки адаптируются к данным условиям культивирования, растут, но не размножаются, они достигают максимальной скорости роста. Абсолютная и удельная скорость роста увеличиваются от нуля до максимально возможных значений.
Абсолютная скорость роста определяется отношением:
V = dx/dt, (1.1)
где V – прирост биомассы или числа клеток;.
х – биомасса или число клеток,
t – время.
Удельная скорость роста определяется по формуле:
µ = (dx/dt) ∙ 1/х, (1.2)
где µ - прирост биомассы е единицу времени на единицу биомассы,
х – начальная биомасса.
Продолжительность лаг-фазы зависит от биологических особенностей бактерий, возраста культуры, количества посевного материала, состава питательной среды, температуры, аэрации, рН и др. Одни бактерии обладают коротким периодом задержки роста, другие длинным. Чем моложе культура, тем период короче. Чем состав питательной среды ближе к тому, в котором выращивали микроорганизмы, тем короче лаг-фаза. Изменения в питательной среде приводят к изменению лаг-фазы, так как необходимо время для синтеза ферментов, либо повышения их активности. Таким образом, факторы задержки роста можно разделить на внешние (состав среды, рН, температура и др.) и внутренние (возраст культуры). Длительность фазы моет быть от нескольких минут до нескольких часов и даже дней. В этой фазе μ = 0.
Логарифмическая , или экспоненциальная , или лог-фаза , характеризуется максимальной скоростью деления бактерий. Экспоненциальный рост популяции описывается уравнением:
Х = Х о ∙ е μ max ∙ t , (1.3)
где Хи Х о - количество клеток (или биомасса) в конце и в начале опыта;
t– время опыта;
е– основание натурального логарифма;
μ max – максимальная удельная скорость роста.
В период логарифмической фазы большинство клеток является физиологически молодыми, биохимически активными, а также наиболее чувствительными к неблагоприятным факторам внешней среды. В этой фазе μ = max.Эта фаза многостадийна, так как в начале ее бактерии растут в среде с избытком субстрата, затем концентрация его понижается, изменяется активность ферментов, возрастает содержание клеточных метаболитов. Кроме того, на рост бактерий оказывают влияние многие факторы: видовые особенности бактерий, характер питательной среды и концентрация ее отдельных компонентов, температура культивирования.
Фаза замедленного роста . Она объединяет две фазы – фазу линейного роста (μ = const) и фазу отрицательного ускорения . Фаза характеризуется в период линейного роста постоянной скоростью прироста биомассы (числа клеток). Затем при переходе в фазу отрицательного ускорения численность делящихся клеток уменьшается. Наступление фазы объясняется количественными изменениями состава питательной среды (потребление питательных веществ, накопление продуктов метаболизма).
Стационарная фаза характеризуется равновесием между погибающими и вновь образующимися клетками. Факторы, лимитирующие рост бактерий в предыдущей фазе, являются причиной возникновения стационарной фазы. Прироста биомассы нет (μ = 0).В этой фазе наблюдается максимальная величина биомассы и максимальная суммарная численность клеток. Эти максимальные величины называются урожаем , или выходом . Одним из ограничивающих факторов является максимальная концентрация клеток в единице объема питательной среды. У разных видов бактерий эта величина значительно варьирует. В стационарной фазе клетки характеризуются несбалансированным ростом (клеточные компоненты синтезируются с различной скоростью), уменьшением интенсивности обменных процессов, более высокой устойчивостью к физическим и химическим воздействиям.
Фаза отмирания (экспоненциальной гибели клеток ) характеризуется уменьшением числа живых клеток, возрастанием гетерогенности популяции (появляются клетки, не воспринимающие краситель, со слабым развитием муреинового слоя и др.). Процесс отмирания превалирует над делением (μ < 0).
Фаза выживания характеризуется наличием отдельных клеток, сохранивших в течение длительного времени жизнеспособность в условиях гибели большинства клеток популяции. Выжившие клетки характеризуются низкой активностью процессов метаболизма, изменением ультраструктуры клеток (мелкозернистая цитоплазма, отсутствие полирибосом и др.). Клетки более устойчивы к неблагоприятным условиям среды.
Таким образом, при стационарном культивировании микробные клетки все время находятся в изменяющихся условиях: сначала имеются в избытке все питательные вещества, затем постепенно наступает их недостаток, затем отравление клеток продуктами метаболизма.
Влияние лимитирующих факторов на скорость роста . Для нормального роста и развития микроорганизмов среда должна содержать необходимые элементы питания, иметь соответствующую рН, температуру и т.д. Факторы, ограничивающие рост культуры, называются лимитирующими . Характерная особенность роста популяции микроорганизмов – зависимость удельной скорости роста от концентрации субстрата. Эта зависимость выражается уравнением Моно , представляющим собой гиперболическую функцию:
μ = μ max ∙ S/(S + K S), (1.4)
где μ – удельная скорость роста;
μ max - максимальная удельная скорость роста;
S – концентрация субстрата;
K S - константанасыщения, численно равная такой концентрации субстрата, которая обеспечивает скорость роста, соответствующую половине значенияμ max .
По мере потребления питательных веществ среда обогащается продуктами обмена, которые также лимитируют рост культуры. Наиболее общий случай влияния концентрации субстрата и продуктов обмена на скорость роста популяции микроорганизмов нашел отражение в модели Н.Д.Иерусалимского:
μ = μ max ∙ S/(S + K S) ∙ К Р / (К Р / + Р), (1.5)
где Р – концентрация продуктов обмена;
К Р - константа, численно равная такой концентрации продуктов обмена, при которой скорость роста замедляется вдвое.
Анализ этого уравнения показывает, что при условии К Р >> Р, когда величиной Р можно пренебречь. скорость роста ограничена только концентрацией субстрата. Если S >> K S , то скорость роста лимитирована накоплением продуктов обмена
Непрерывное культивирование. Если в емкость, где находится бактериальная популяция, непрерывно подавать свежую питательную среду и одновременно с такой же скоростью выводить культуральную жидкость, содержащую бактериальные клетки и продукты метаболизма, то получается непрерывное культивирование. Регулируя скорость проточной среды, можно управлять ростом бактериальной популяции, например, удлинять логарифмическую или стационарную фазу на любое необходимое время. Непрерывное культивирование осуществляется в специальных приборах - хемостатах и турбидостатах.
Хемостаты . Рост бактерий регулируется концентрацией субстрата. Поддерживая постоянную концентрацию одного из необходимых субстратов (источник азота или углерода), путем регулирования скорости протока среды, можно сбалансировать скорость роста культуры. Скорость изменения величины биомассы клеток в хемостате равна разности между скоростью прироста биомассы и скоростью выноса ее из культиватора. Плотность популяции остается постоянной, если μ=D (удельная скорость роста равна коэффициенту разбавления), т.е. потеря клеток в результате вымывания и прирост их в результате размножения уравновешивается.
Турбидостаты .Принцип работы основан на регулировании скорости потока среды плотностью популяции. Плотность популяции контролируется фотоэлементом, соединенным с реле, регулирующим подачу среды. Когда плотность популяции достигает заданного уровня, реле срабатывает и в культиватор поступает свежая среде. В результате концентрация клеток уменьшается до определенного уровня и затем автоматически отключается подача среды.
Турбидостатный контроль может быть основан на других метолах определения биомассы, либо продуктов, образующихся в процессе роста бактерий (например, рН-статный способ управления скорости потока, использование оксистата– управление скоростью потока по скорости потребления кислорода и др.).
Непрерывное культивирование микроорганизмов используется для изучения их физиологии, биохимии, генетики и др., а также широко используется в микробиологической промышленности.
Синхронное культивирование. Синхронные культуры – это культуры, в которых некоторое время все клетки делятся одновременно (синхронно) за счет одинаковой готовности к делению всех особей. Синхронизация достигается физическими и химико-биологическими методами. Физические методы - это температурное воздействие, дифференциальное центрифугирование или дифференциальное фильтрование и др. Химико-биологические методы: вынужденное голодание бактерий, выращивание бактерий на неполноценных средах с последующим переносом их в полноценные среды. Синхронные культуры используются для генетических и цитологических исследований, для изучения синтеза отдельных клеточных компонентов в процессе деления бактерий.
Рост бактерий происходит в результате множества взаимосвязанных биохимических реакций , осуществляющих синтез клеточного материала. У бактерий различают индивидуальный рост бактериальной клетки и рост бактерий в популяции.
Об индивидуальном росте судят по увеличению размеров отдельных особей. Скорость роста зависит от внешних условий и физиологического состояния самой клетки. При постоянных условиях рост осуществляется с постоянной скоростью. Палочковидные бактерии растут преимущественно в направлении длинной оси, кокки растут равномерно во всех направлениях. В промежутке между клеточными делениями бактерии имеют большие размеры, чем сразу после деления.
Размножение бактерий
Наиболее часто бактерии размножаются путем бинарного деления, когда из одной клетки образуется две, каждая из которых вновь делится. Процессу деления всегда предшествует репликация (удвоение) ДНК . Существует два типа деления - деление перетяжкой (перешнуровывание) и с помощью поперечной перегородки (рисунок А.7) .
Деление перетяжкой (констрикция) сопровождается сужением клетки в месте ее деления, и в этом процессе принимают участие все слои клеточных оболочек. Выпячивание оболочек внутрь клетки все более ее сужает и, наконец, делит на две. Это деление присуще грамотрицательным бактериям . Деление с образованием поперечной перегородки присуще грамположительным бактериям. Однако у некоторых групп бактерий отмечена смена способов деления (тионовые бактерии, микобактерии). У шаровидных бактерий может образовываться несколько поперечных перегородок (тетракокки, сарцины). Почкование убактерий является разновидностью бинарного деления. Этот способ размножения присущ бактериям, имеющим диморфные или полиморфные клеточные циклы. Почкующимся бактериям присуща полярность клеток. Некоторые бактерии размножаются с помощью экзоспор (но не эндоспор!), некоторые - фрагментами гиф (актиномицеты). У некоторых бактерий имеются половые ворсинки, или F-пили.
Период от деления до деления называется клеточным циклом . Различают несколько типов вегетативного клеточного цикла: мономорфный - образуется только один морфологический тип клеток (например, бациллы), диморфный - два морфологических типа клеток, полиморфный - несколько (актиномицеты). При диморфном и полиморфном циклах различают дочерние и материнские клетки.
Бактерии характеризуются высокой скоростью размножения. Например, в благоприятных условиях кишечная палочка делится каждые 20-30 мин, за сутки это дает 2 72 , т.е. 72 поколения. В условиях, исключающих гибель, эта биомасса составит 4720 т. Скорость размножения зависит от факторов внешней среды (температуры, условия питания, влажность, реакция среды и др.) и от видовых особенностей бактерий. Высокая скорость размножения бактерий обеспечивает их сохранение на земле даже в условиях массовой гибели. Сохранившиеся отдельные клетки размножаются и вновь дают поколение.
Рост бактерий в популяции. Популяция (фр. population - население) - это совокупность бактерий одного вида (чистая культура) или разных видов (смешанная ассоциация), развивающихся в ограниченном пространстве (например, в питательной среде). В бактериальной популяции постоянно происходит рост, размножение и отмирание клеток. Культивирование микроорганизмов в искусственных условиях бывает периодическим, непрерывным и синхронным.
Периодическое (стационарное) культивирование происходит без притока и оттока питательной среды. Оно характеризуется классической кривой роста микроорганизмов, в которой выделяют отдельные фазы роста бактериальной популяции, отражающие общую закономерность роста и размножения клеток.
Лаг-фаза (англ. lag - отставание) начинается с момента посева бактерий в свежую питательную среду. Клетки адаптируются к данным условиям культивирования, растут, но не размножаются, они достигают максимальной скорости роста. Абсолютная и удельная скорость роста увеличиваются от нуля до максимально возможных значений.
Абсолютная скорость роста определяется отношением:
V = dx/dt , (1.1)
где V - прирост биомассы или числа клеток, выражается в массовых единицах, числе клеток или в условных единицах в единицу времени.
х - биомасса или число клеток;
t - время.
Удельная скорость роста определяется по формуле:
µ = (dx/dt) ? 1/х , (1.2)
где µ - прирост биомассы е единицу времени на единицу биомассы,
х - начальная биомасса.
Продолжительность лаг-фазы зависит от биологических особенностей бактерий, возраста культуры, количества посевного материала, состава питательной среды, температуры, аэрации, рН и др. Одни бактерии обладают коротким периодом задержки роста, другие длинным. Чем моложе культура, тем период короче. Чем состав питательной среды ближе к тому, в котором выращивали микроорганизмы, тем короче лаг-фаза. Изменения в питательной среде приводят к изменению лаг-фазы, так как необходимо время для синтеза ферментов, либо повышения их активности. Таким образом, факторы задержки роста можно разделить на внешние (состав среды, рН, температура и др.) и внутренние (возраст культуры). Длительность фазы моет быть от нескольких минут до нескольких часов и даже дней. В этой фазе μ = 0.
Лог-фаза (логарифмическая , или экспоненциальная ) характеризуется максимальной скоростью деления бактерий. Общее количество бактерий определяется по формуле:
N = N 0 ?2 n , (1.3)
где N и N 0 - общее количество клеток в конце фазы и в начале фазы соответственно;
n - число поколений, или генераций.
В микробиологической практике для выражения общего числа микробных клеток чаще всего пользуются не абсолютными числами (так как они достигают огромных величин), а их логарифмами. Прсле логарифмирования уравнения (1.3): lg N = lg N 0 + n?lg2, n?lg2 = lg N - lg N 0 , отсюда число поколений равно: n = (lg N - lg N 0)/ lg2
Скорость размножения одной клетки, или период генерации:
g = t/n , (1.4)
где t - время;
n - число поколений;
g - период генерации.
Значит: g = t ?lg 2 / (lg N - lg N 0) (1.5)
Приведенные уравнения основаны на предположении, что в лог-фазе все 100% клеток жизнеспособны. Однако экспериментально установлено, что около 20% клеток даже в эту фазу отмирает, поэтому в приведенные формулы вносится поправка - вместо 2 берется 1,6.
Экспоненциальный рост популяции описывается уравнением:
Х = Х 0 ? е μ max ? t , (1.6)
где Х и Х 0 - количество клеток (или биомасса) в конце и в начале опыта соответственно;
t - время опыта;
е - основание натурального логарифма;
μ max - максимальная удельная скорость роста.
В период логарифмической фазы большинство клеток является физиологически молодыми, биохимически активными, а также наиболее чувствительными к неблагоприятным факторам внешней среды. В этой фазе μ = max.
Фаза замедленного роста . Она объединяет две фазы - фазу линейного роста (μ = const) и фазу отрицательного ускорения . Фаза характеризуется в период линейного роста постоянной скоростью прироста биомассы (числа клеток). Затем при переходе в фазу отрицательного ускорения численность делящихся клеток уменьшается. Наступление фазы объясняется количественными изменениями состава питательной среды (потребление питательных веществ, накопление продуктов метаболизма).
Стационарная фаза характеризуется равновесием между погибающими и вновь образующимися клетками. Факторы, лимитирующие рост бактерий в предыдущей фазе, являются причиной возникновения стационарной фазы. Прироста биомассы нет (μ = 0) . В этой фазе наблюдается максимальная величина биомассы и максимальная суммарная численность клеток. Эти максимальные величины называются урожаем , или выходом .
Фаза отмирания (экспоненциальной гибели клеток ) характеризуется уменьшением числа живых клеток, возрастанием гетерогенности популяции (появляются клетки, не воспринимающие краситель, со слабым развитием муреинового слоя и др.). Процесс отмирания превалирует над делением (μ < 0).
Фаза выживания характеризуется наличием отдельных клеток, сохранивших в течение длительного времени жизнеспособность в условиях гибели большинства клеток популяции. Выжившие клетки характеризуются низкой активностью процессов метаболизма, изменением ультраструктуры клеток (мелкозернистая цитоплазма, отсутствие полирибосом и др.). Клетки более устойчивы к неблагоприятным условиям среды.
Таким образом, при стационарном культивировании микробные клетки все время находятся в изменяющихся условиях: сначала имеются в избытке все питательные вещества, затем постепенно наступает их недостаток, затем отравление клеток продуктами метаболизма.
Влияние лимитирующих факторов на скорость роста . Для нормального роста и развития микроорганизмов среда должна содержать необходимые элементы питания, иметь соответствующую рН, температуру и т.д. Факторы, ограничивающие рост культуры, называются лимитирующими . Характерная особенность роста популяции микроорганизмов - зависимость удельной скорости роста от концентрации субстрата. Эта зависимость выражается уравнением Моно , представляющим собой гиперболическую функцию:
μ = μ max ? S/(S + K S) , (1.7)
где μ - удельная скорость роста;
μ max - максимальная удельная скорость роста;
S - концентрация субстрата;
K S - константанасыщения, численно равная такой концентрации субстрата, которая обеспечивает скорость роста, соответствующую половине значенияμ max .
По мере потребления питательных веществ среда обогащается продуктами обмена, которые также лимитируют рост культуры. Наиболее общий случай влияния концентрации субстрата и продуктов обмена на скорость роста популяции микроорганизмов нашел отражение в модели Н.Д. Иерусалимского:
μ = μ max ? S/(S + K S) ? К Р / (К Р + Р) , (1.8)
где Р - концентрация продуктов обмена;
К Р - константа, численно равная такой концентрации продуктов обмена, при которой скорость роста замедляется вдвое.
Анализ этого уравнения показывает, что при условии К Р >> Р, когда величиной Р можно пренебречь. скорость роста ограничена только концентрацией субстрата. Если S >> K S , то скорость роста лимитирована накоплением продуктов обмена.
Непрерывное культивирование . Если в емкость, где находится бактериальная популяция, непрерывно подавать свежую питательную среду и одновременно с такой же скоростью выводить культуральную жидкость, содержащую бактериальные клетки и продукты метаболизма, то получается непрерывное культивирование. Регулируя скорость проточной среды, можно управлять ростом бактериальной популяции, например, удлинять логарифмическую или стационарную фазу на любое необходимое время. Непрерывное культивирование осуществляется в специальных приборах - хемостатах и турбидостатах. Непрерывное культивирование микроорганизмов используется для изучения их физиологии, биохимии, генетики и др., а также широко используется в микробиологической промышленности.
Синхронные культуры - это культуры, в которых некоторое время все клетки делятся одновременно (синхронно) за счет одинаковой готовности к делению всех особей. Синхронизация достигается физическими и химико-биологическими методами. К физическим методам относится температурное воздействие, дифференциальное центрифугирование или дифференциальное фильтрование, химико-биологическим - вынужденное голодание бактерий, выращивание бактерий на неполноценных средах с последующим переносом их в полноценные среды. Синхронные культуры используются для генетических и цитологических исследований, для изучения синтеза отдельных клеточных компонентов в процессе деления бактерий.
Размножение бактерий путем деления — самый распространенный метод увеличения численности микробной популяции. После деления происходит рост бактерий до исходного размера, для чего необходимы определенные вещества (факторы роста).
Способы размножения бактерий различны, но для большинства их видов присуща форма бесполового размножения способом деления. Способом почкования бактерии размножаются исключительно редко. Половое размножение бактерий присутствует в примитивной форме.
Рис. 1. На фото бактериальная клетка в стадии деления.
Генетический аппарат бактерий
Генетический аппарат бактерий представлен единственной ДНК — хромосомой. ДНК замкнута в кольцо. Хромосома локализована в нуклеотиде, не имеющем мембраны. В бактериальной клетке имеются плазмиды.
Нуклеоид
Нуклеоид является аналогом ядра. Он расположен в центре клетки. В нем локализована ДНК — носитель наследственной информации в свернутом виде. Раскрученная ДНК достигает в длину 1 мм. Ядерное вещество бактериальной клетки не имеет мембраны, ядрышка и набора хромосом, не делится митозом. Перед делением нуклеотид удваивается. Во время деления число нуклеотидов увеличивается до 4-х.

Рис. 2. На фото бактериальная клетка на срезе. В центральной части виден нуклеотид.
Плазмиды
Плазмиды представляют собой автономные молекулы свернутые в кольцо двунитевой ДНК. Их масса значительно меньше массы нуклеотида. Несмотря на то, что в ДНК плазмид закодирована наследственная информация, они не являются жизненно важными и необходимыми для бактериальной клетки.

Рис. 3. На фото бактериальная плазмида.
Этапы деления
После достижения определенных размеров, присущих взрослой клетке, запускаются механизмы деления.
Репликация ДНК
Репликация ДНК предшествует клеточному делению. Мезосомы (складки цитоплазматической мембраны) удерживают ДНК до тех пор, пока процесс деления (репликации) не завершится.
Репликация ДНК осуществляется с помощью ферментов ДНК-полимеразами. При репликации водородные связи в 2-х спиральной ДНК разрываются, в результате чего из одной ДНК образуются две дочерние односпиральные. В последующем, когда дочерние ДНК заняли свое место в разделенных дочерних клетках, происходит их восстановление.
Как только репликация ДНК завершилась, в результате синтеза появляется перетяжка, разделяющая клетку пополам. Вначале делению подвергается нуклеотид, затем цитоплазма. Синтез клеточной стенки завершает деление.
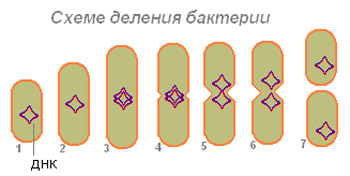
Рис. 4. Схема деления бактериальной клетки.
Обмен участками ДНК
У сенной палочки процесс репликации ДНК завершается обменом участками 2-х ДНК.
После деления клетки образуется перемычка, по которой ДНК одной клетки переходит в другую. Далее обе ДНК сплетаются. Некоторые отрезки обоих ДНК слипаются. В местах слипания происходит обмен отрезками ДНК. Одна из ДНК по перемычке уходит обратно в первую клетку.

Рис. 5. Вариант обмена ДНК у сенной палочки.
Типы делений бактериальных клеток
Если клеточное деление опережает процесс разделения, то образуются многоклеточные палочки и кокки.
При синхронном клеточном делении образуются две полноценные дочерние клетки.
Если нуклеотид делится быстрее самой клетки, то образуются многонуклеотидные бактерии.
Способы разделения бактерий
Деление с помощью разламывания
Деление с помощью разламывания характерно для сибиреязвенных бацилл. В результате такого деления клетки переламываются в местах сочленения, разрывая цитоплазматические мостики. Далее отталкиваются друг от друга, образуя цепочки.
Скользящее разделение
При скользящем разделении после деления клетка обосабливается и как бы скользит по поверхности другой клетки. Данный способ разделения характерен для некоторых форм эшерихий.
Секущееся разделение
При секущемся разделении одна из разделившихся клеток свободным концом описывает дугу круга, центром которого является точка ее контакта с другой клеткой, образуя римскую пятерку или клинопись (коринебактерии дифтерии, листерии).

Рис. 6. На фото бактерии палочковидной формы, образующие цепочки (сибиреязвенные палочки).

Рис. 7. На фото скользящий способ разделения кишечных палочек.

Рис. 8. Секущийся способ разделения коринебактерий.
Вид скоплений бактерий после деления
Скопления делящихся клеток имеют разнообразную форму, которая зависит от направления плоскости деления.
Шаровидные бактерии располагаются по одному, по двое (диплококки), пакетами, цепочками или как гроздья винограда. Палочковидные бактерии — цепочками.
Спиралевидные бактерии — хаотично.

Рис. 9. На фото микрококки. Они круглые, гладкие, имеют белую, желтую и красную окраску. В природе микрококки распространены повсеместно. Живут в разных полостях человеческого организма.

Рис. 10. На фото бактерии диплококки — Streptococcus pneumoniae.

Рис. 11. На фото бактерии сарцины. Кокковидные бактерии соединяются в пакеты.

Рис. 12. На фото бактерии стрептококки (от греческого «стрептос» — цепочка). Располагаются цепочками. Являются возбудителями целого ряда заболеваний.

Рис. 13. На фото бактерии «золотистые» стафилококки. Располагаются, как «гроздья винограда». Скопления имеют золотистую окраску. Являются возбудителями целого ряда заболеваний.

Рис. 14. На фото извитые бактерии лептоспиры — возбудители многих заболеваний.

Рис. 15. На фото палочковидные бактерии рода Vibrio.
Скорость деления бактерий
Скорость деления бактерий крайне высока. В среднем одна бактериальная клетка делится каждые 20 минут. В течение только одних суток одна клетка образует 72 поколения потомства. Микобактерии туберкулеза делятся медленно. Весь процесс деления занимает у них около 14 часов.

Рис. 16. На фото отображен процесс деления клетки стрептококка.
Половое размножение бактерий
В 1946 году учеными было обнаружено половое размножение в примитивной форме. При этом гаметы (мужские и женские половые клетки) не образуются, однако некоторые клетки обмениваются генетическим материалом (генетическая рекомбинация ).
Передача генов осуществляется в результате конъюгации — однонаправленного переноса части генетической информации в виде плазмид при контакте бактериальных клеток.
Плазмиды представляют собой молекулы ДНК небольшого размера. Они не связаны с геномом хромосом и способны удваиваться автономно. В плазмидах содержаться гены, которые повышают устойчивость бактериальных клеток к неблагоприятным условиям внешней среды. Бактерии часто передают эти гены друг другу. Отмечается так же передача генной информации бактериям другого вида.
При отсутствии истинного полового процесса именно конъюгация играет огромную роль при обмене полезными признаками. Так передается способность бактерий проявлять лекарственную устойчивость. Для человечества особо опасным является передача устойчивости к антибиотикам между болезнетворными популяциями.

Рис. 17. На фото момент конъюгации двух кишечных палочек.
Фазы развития бактериальной популяции
При посевах на питательную среду развитие бактериальной популяции проходит несколько фаз.
Исходная фаза
Исходная фаза — это период от момента посева до их роста. В среднем исходная фаза длится 1 — 2 часа.
Фаза задержки размножения
Это фаза интенсивного роста бактерий. Ее длительность составляет около 2-х часов. Она зависит от возраста культуры, периода приспособления, качества питательной среды и др.
Логарифмическая фаза
В эту фазу отмечается пик скорости размножения и увеличения бактериальной популяции. Ее длительность составляет 5 — 6 часов.
Фаза отрицательного ускорения
В эту фазу отмечается спад скорости размножения, уменьшается количество делящихся и увеличивается число погибших бактерий. Причина отрицательного ускорения — истощение питательной среды. Ее длительность составляет около 2-х часов.
Стационарная фаза максимума
В стационарную фазу отмечается равное количество погибших и вновь образованных особей. Ее длительность составляет около 2-х часов.
Фаза ускорения гибели
В эту фазу прогрессивно нарастает количество погибших клеток. Ее длительность составляет около 3-х часов.
Фаза логарифмической гибели
В эту фазу клетки бактерий отмирают с постоянной скоростью. Ее длительность составляет около 5-и часов.
Фаза уменьшения скорости отмирания
В эту фазу оставшиеся живыми клетки бактерий переходят в состояние покоя.

Рис. 18. На рисунке отображена кривая роста бактериальной популяции.

Рис. 19. На фото колонии синегнойной палочки сине-зеленого цвета, колонии микрококков желтого цвета, колонии Bacterium prodigiosum кроваво-красного цвета и колонии Bacteroides niger черного цвета.

Рис. 20. На фото колонии бактерий. Каждая колония — потомство одной-единственной клетки. В колонии число клеток исчисляется миллионами. вырастает колония за 1 — 3 суток.
Деление магниточувствительных бактерий
В 1970-х годах были открыты бактерии, обитающие в морях, которые обладали чувством магнетизма. Магнетизм позволяет этим удивительным существам ориентироваться по линиям магнитного поля Земли и находить серу, кислород и другие, так необходимые ей вещества. Их «компас» представлен магнитосомами, которые состоят из магнита. При делении магниточувствительные бактерии делят свой компас. При этом перетяжки при делении становится явно недостаточно, поэтому бактериальная клетка сгибается и делает резкий перелом.

Рис. 21. На фото момент деления магниточувствительной бактерии.
Рост бактерий
Вначале деления бактериальной клетки две молекулы ДНК расходятся в разные концы клетки. Далее клетка делится на две равноценные части, которые отделяются друг от друга и увеличиваются до исходного размера. Скорость деления многих бактерий составляет в среднем 20 — 30 минут. В течение только одних суток одна клетка образует 72 поколения потомства.
Масса клеток в процессе роста и развития быстро поглощает питательные вещества из окружающей среды. Этому способствуют благоприятные факторы внешней среды — температурный режим, достаточное количество питательных веществ, необходимая pH среды. Для клеток аэробов необходим кислород. Для анаэробов он представляет опасность. Однако безграничное размножение бактерий в природе не происходит. Солнечный свет, сухой воздух, недостаток пищи, высокая температура окружающей среды и другие факторы губительно действуют на бактериальную клетку.

Рис. 22. На фото момент деления клетки.
Факторы роста
Для роста бактерий необходимы определенные вещества (факторы роста), часть из которых синтезируется самой клеткой, часть поступает из окружающей среды. Потребность в факторах роста у всех бактерий разная.
Потребность в факторах роста является постоянным признаком, что позволяет использовать его для идентификации бактерий, подготовке питательных сред и использовать в биотехнологии.
Факторы роста бактерий (бактериальные витамины) — химические элементы, большинством из которых являются водорастворимые витамины группы В. В эту группу входят так же гемин, холин, пуриновые и пиримидиновые основания и другие аминокислоты. При отсутствии факторов роста наступает бактериостаз.
Бактерии используют факторы роста в минимальных количествах и в неизменном виде. Ряд химических веществ этой группы входят в состав клеточных ферментов.

Рис. 23. На фото момент деления палочковидной бактерии.
Важнейшие бактериальные факторы роста
- Витамин В1 (тиамин) . Принимает участие в углеводном обмене.
- Витамин В2» (рибофлавин) . Принимает участие в окислительно-восстановительных реакциях.
- Пантотеновая кислота является составной частью кофермента А.
- Витамин В6 (пиридоксин) . Принимает участие в обмене аминокислот.
- Витамины В12 (кобаламины — вещества, содержащие кобальт). Принимают активное участие в синтезе нуклеотидов.
- Фолиевая кислота . Некоторые ее производные входят в состав ферментов, катализирующих процессы синтеза пуриновых и пиримидиновых оснований, а также некоторых аминокислот.
- Биотин . Участвует в азотистом обмене, а также катализирует синтез ненасыщенных жирных кислот.
- Витамин РР (никотиновая кислота). Участвует в окислительно-восстановительных реакциях, образовании ферментов и обмене липидов и углеводов.
- Витамин Н (парааминобензойная кислота). Является фактором роста многих бактерий, в том числе населяющих кишечник человека. Из парааминобензойной кислоты синтезируется фолиевая кислота.
- Гемин . Является составной частью некоторых ферментов, которые принимают участие в реакциях окислениях.
- Холин . Принимает участие в реакциях синтеза липидов клеточной стенки. Является поставщиком метильной группы при синтезе аминокислот.
- Пуриновые и пиримидиновые основания (аденин, гуанин, ксантин, гипоксантин, цитозин, тимин и урацил). Вещества необходимы главным образом в качестве компонентов нуклеиновых кислот.
- Аминокислоты . Эти вещества являются составляющими белков клетки.
Потребность в факторах роста некоторых бактерий
Ауксотрофы для обеспечения жизнедеятельности нуждаются в поступлении химических веществ из вне. Например, клостридии не способны синтезировать лецитин и тирозин. Стафилококки нуждаются в поступлении лецитина и аргинина. Стрептококки нуждаются в поступлении жирных кислот — компонентов фосфолипидов. Коринебактерии и шигеллы нуждаются в поступлении никотиновой кислоты. Золотистые стафилококки, пневмококки и бруцеллы нуждаются в поступлении витамина В1. Стрептококки и бациллы столбняка — в пантотеновой кислоте.
Прототрофы самостоятельно синтезируют необходимые вещества.

Рис. 24. Разные условия окружающей среды по-разному влияют на рост колоний бактерий. Слева — стабильный рост в виде медленно расширяющегося круга. Справа — быстрый рост в виде «побегов».
Изучение потребности бактерий в факторах роста позволяет ученым получать большую микробную массу, так необходимую при изготовлении антимикробных препаратов, сывороток и вакцин.
Подробно о бактерияx читай в статьях:
Размножение бактерий является механизмом повышения числа микробной популяции. Деление бактерий — основной способ размножения. После деления бактерии должны достигнуть размеров взрослых особей. Рост бактерий происходит путем быстрого поглощения питательных веществ их окружающей среды. Для роста необходимы определенные вещества (факторы роста), часть из которых синтезирует сама бактериальная клетка, часть поступает из окружающей среды.
Изучая рост и размножение бактерий, ученые постоянно открывают полезные свойства микроорганизмов, использование которых в повседневной жизни и на производстве ограничивается только их свойствами.
Жизнедеятельность бактерий характеризуется ростом -- формированием структурно-функциональных компонентов клетки и увеличением самой бактериальной клетки, а также размножением -- самовоспроизведением, приводящим к увеличению количества бактериальных клеток в популяции.
Бактерии размножаются путем бинарного деления пополам, реже путем почкования. Актиномицеты, как и грибы, могут размножаться спорами. Актиномицеты, являясь ветвящимися бактериями, размножаются путем фрагментации нитевидных клеток. Грамположительные бактерии делятся путем врастания синтезирующихся перегородок деления внутрь клетки, а грамотрицательные -- путем перетяжки, в результате образования гантелевид-ных фигур, из которых образуются две одинаковые клетки.
Делению клеток предшествует репликация бактериальной хромосомы по полуконсервативному типу (двуспиральная цепь ДНК раскрывается и каждая нить достраивается комплементарной нитью), приводящая к удвоению молекул ДНК бактериального ядра -- нуклеоида.
Репликация ДНК происходит в три этапа: инициация, элонгация, или рост цепи, и терминация.
Размножение бактерий в жидкой питательной среде. Бактерии, засеянные в определенный, не изменяющийся объем питательной среды, размножаясь, потребляют питательные элементы, что приводит в дальнейшем к истощению питательной среды и прекращению роста бактерий. Культивирование бактерий в такой системе называют периодическим культивированием, а культуру -- периодической. Если же условия культивирования поддерживаются путем непрерывной подачи свежей питательной среды и оттока такого же объема культуральной жидкости, то такое культивирование называется непрерывным, а культура -- непрерывной.
При выращивании бактерий на жидкой питательной среде наблюдается придонный, диффузный или поверхностный (в виде пленки) рост культуры. Рост периодической культуры бактерий, выращиваемых на жидкой питательной среде, подразделяют на несколько фаз, или периодов :
- 1. лаг-фаза;
- 2. фаза логарифмического роста;
- 3. фаза стационарного роста, или максимальной концентрации
бактерий;
4. фаза гибели бактерий.
Эти фазы можно изобразить графически в виде отрезков кривой размножения бактерий, отражающей зависимость логарифма числа живых клеток от времени их культивирования.
Лаг-фаза -- период между посевом бактерий и началом размножения. Продолжительность лаг-фазы в среднем 4--5 ч. Бактерии при этом увеличиваются в размерах и готовятся к делению; нарастает количество нуклеиновых кислот, белка и других компонентов.
Фаза логарифмического (экспоненциального) роста является периодом интенсивного деления бактерий. Продолжительность ее около 5-- 6 ч. При оптимальных условиях роста бактерии могут делиться каждые 20--40 мин. Во время этой фазы бактерии наиболее ранимы, что объясняется высокой чувствительностью компонентов метаболизма интенсивно растущей клетки к ингибиторам синтеза белка, нуклеиновых кислот и др.
Затем наступает фаза стационарного роста , при которой количество жизнеспособных клеток остается без изменений, составляя максимальный уровень (М-концентрация). Ее продолжительность выражается в часах и колеблется в зависимости от вида бактерий, их особенностей и культивирования.
Завершает процесс роста бактерий фаза гибели , характеризующаяся отмиранием бактерий в условиях истощения источников питательной среды и накопления в ней продуктов метаболизма бактерий. Продолжительность ее колеблется от 10 ч до нескольких недель. Интенсивность роста и размножения бактерий зависит от многих факторов, в том числе оптимального состава питательной среды, окислительно-восстановительного потенциала, рН, температуры и др.
Размножение бактерий на плотной питательной среде. Бактерии, растущие на плотных питательных средах, образуют изолированные колонии округлой формы с ровными или неровными краями (S- и R-формы), различной консистенции и цвета, зависящего от пигмента бактерий.
Пигменты, растворимые в воде, диффундируют в питательную среду и окрашивают её. Другая группа пигментов нерастворима в воде, но растворима в органических растворителях. И, наконец, существуют пигменты, не растворимые ни в воде, ни в органических соединениях.
Наиболее распространены среди микроорганизмов такие пигменты, как каротины, ксантофиллы и меланины. Меланины являются нерастворимыми пигментами черного, коричневого или красного цвета, синтезирующимися из фенольных соединений. Меланины наряду с каталазой, супероксидцисмутазой и пероксидазами защищают микроорганизмы от воздействия токсичных перекисных радикалов кислорода. Многие пигменты обладают антимикробным, антибиотикоподобным действием.